|
|
|
Free Neuropathology 4:8 (2023) |
|
Review |
|
Neurodevelopmental disorders: 2023 update |
|
Paulina Carriba1,3, Nicola Lorenzón1, Mara Dierssen1,2,3 |
|
|
Corresponding author: |
|
Submitted: 28 February 2023 |
|
Keywords: Centrosomes, CLIP: Caudal late interneuron progenitor, Human organoids, MCD: malformations of cortical development, Stress granules, SUDC: sudden unexplained death in childhood |
|
Abstract Several advances in the field of neurodevelopmental diseases (NDDs) have been reported by 2022. Of course, NDDs comprise a diverse group of disorders, most of which with different aetiologies. However, owing to the development and consolidation of technological approaches, such as proteomics and RNA-sequencing, and to the improvement of brain organoids along with the introduction of artificial intelligence (AI) for biodata analysis, in 2022 new aetiological mechanisms for some NDDs have been proposed. Here, we present hints of some of these findings. For instance, centrioles regulate neuronal migration and could be behind the aetiology of periventricular heterotopia; also, the accumulation of misfolded proteins could explain the neurological effects in COVID-19 patients; and, autism spectrum disorders (ASD) could be the expression of altered cortical arealization. We also cover other interesting aspects as the description of a new NDD characterized by deregulation of genes involved in stress granule (SG) assemblies, or the description of a newly discovered neural progenitor that explains the different phenotypes of tumours and cortical tubers in tuberous sclerosis complex (TSC) disease; and how it is possible to decipher the aetiology of sudden unexplained death in childhood (SUDC) or improve the diagnosis of cortical malformations using formalin-fixed paraffin-embedded samples. |
|
Abbreviations AD – Alzheimer’s disease; ADHD - attention deficit/hyperactivity disorder; AI – artificial intelligence; ASD - autism spectrum disorder; BCs – balloon cells; CA – cornu ammonis; CD - cluster of differentiation; CGE - caudal ganglionic eminence; CLIP - caudal late interneuron progenitor; CNS – central nervous system; DG – dentate gyrus; EGFR - epidermal growth factor receptor; EIF2 - Eukaryotic Initiation Factor 2; FCD – focal cortical dysplasia; FFPE - formalin-fixed paraffin-embedded; FS - febrile seizure; GCs - giant cells; GO – gene ontology; HAND – HIV-associated neurocognitive disorder; hCO - human cortical organoids; Het – heterozygous; HIV – human immunodeficiency virus; ID - intellectual disability; iPSCs - induced pluripotent stem cells; KO – knockout; LOH - loss of heterozygosity; MCDs – malformations of cortical development; mTOR - mammalian target of rapamycin; NSC – neural stem cells; ORF – open reading frame; PH – periventricular heterotopia; PRPF6 - pre-mRNA processing factor 6; RNA-seq – RNA sequencing; SARS-CoV-2 - severe acute respiratory syndrome coronavirus 2; SGs - stress granules; SUDC - sudden unexplained death in childhood; SUDEP - sudden unexpected death in epilepsy; t-hCO – transplanted human cortical organoids; TS – Timothy syndrome; TSC - tuberous sclerosis complex; WT - wild-type. Introduction For this new collection of the most relevant findings in neurodevelopmental disorders that appeared in 2022, our selection tried to encompass a wide range of aspects that we think could be of interest to neuropathologists. The topics chosen are:
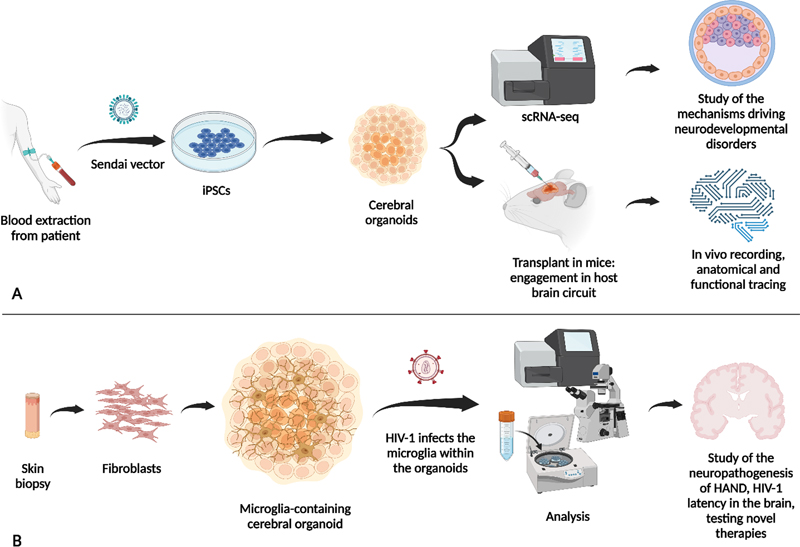
Along these subject matters we aim to discuss advances in different NDDs, from brain malformations or classical neurodevelopmental conditions to more general aspects that a neuropathologist might face, such as paediatric neurological alterations associated with COVID-19 or HIV-associated neurocognitive disorder (HAND) in children. We also selected relevant findings regarding how formalin-fixed paraffin-embedded (FFPE) samples, the major form of stored brain samples, could be used for studying neurodevelopmental disorders, and how the use of artificial intelligence (AI) can improve the diagnosis of cortical malformations. Some of the selected topics also provide new mechanistic insights, such as the newly discovered neural progenitor CLIP, which explains the divergent phenotypes in tuberous sclerosis complex (TSC) pathology. Indeed, one of the most remarkable topics of 2022, which has acquired increasing relevance in recent years, is the importance of the spatial and temporal regulation of brain development. The coordinated and orchestrated series of cellular processes controlled by fine-tuned sets of genetic programs during neurodevelopment leads to immense cell diversity, with different features depending on their final fate, localisation, and properties distinctive from the cells from which they developed. Those cells will be part of circuits that are adjusted, readjusted and refined by intrinsic and extrinsic signals (Rubenstein and Rakic, 1999; Miyata et al., 2010; Kwan et al., 2012; Greig et al., 2013; Wamsley et al., 2018; Di Bella et al., 2021; Bonnefont and Vanderhaeghen, 2021) in a precise spatial-temporal manner. Within this choreographic arrangement, a single out-of-tune event in time or space may represent the inception of a neurodevelopmental pathological condition. Finally, human-derived organoids continue to be a promising in vitro tool for modelling human physiological and pathological development (Figure 1). In the last year, those systems gained popularity thanks to specific improvements in successfully modelling neurodevelopmental disorders, allowing the study of human neuronal function in an in vivo context.
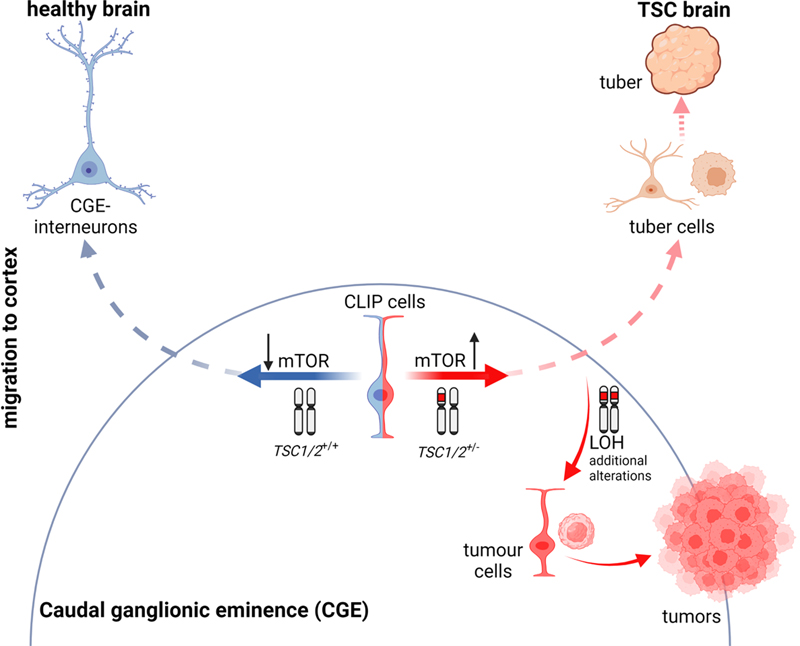
Figure 1. Schematic representation of some uses of human-derived organoids presented in this update. A) Cerebral organoids derived from iPSCs can be used for single-cell RNA-sequencing (scRNAseq) to determine expression patterns for the study of mechanisms driving neurodevelopmental disorders. Here presented in Neurodevelopmental diseases and the proper space and time sequential events during brain neurodevelopment; CLIP, a newly discovered interneuron progenitor, explains the divergent phenotype in tuberous sclerosis complex disorder; and in Transcriptomic dysregulation in ASD occurs across the whole cerebral cortex and follows a regional gradient. Also, cerebral organoids can be transplanted into a host brain circuit to study the functionality of human organoids, as introduced in the in vivo platform for the study of human neurodevelopmental diseases. B) New engineering organoids containing microglia have been developed last year to study novel mechanisms explaining COVID-19 neurological anomalies. 1. Neurodevelopmental diseases and the proper space and time sequential events during brain neurodevelopment Although it has been suggested that out-of-tune events at specific time points or specific brain regions are crucial for understanding neurodevelopmental pathological conditions, few examples have established a concrete cellular process in which such time- and place-specific effects could be disentangled. O’Neill et al. reported time-dependent dysregulation of the centrosome interactome at specific neuronal differentiation stages, which allowed studying the aetiology of neurodevelopmental diseases (O’Neill et al., 2022). Centrosomes, as anchor structures for the cell cytoskeleton, are involved in a number of cell functions, including mitosis and cell migration (Wilsch-Bräuninger and Huttner, 2021; Gönczy and Hatzopoulos, 2019; Vineethakumari and Lüders, 2022; Delgehyr et al., 2005; Piel et al., 2000). To prove their time-dependent hypothesis, the authors derived neural stem cells (NSC) [15 days in culture], and differentiated neurons [40 days in culture] to forebrain identity, using human induced pluripotent stem cell (iPSC) lines. At these two stages, mass spectrometry (MS) of centrosome-associated proteins revealed large cell type-specificity, with around 60% of the neural centrosome proteins not being detected in the centrosome of other cell types. Gene Ontology (GO) categorization indicated that, as expected, NSC centrosome-associated proteins are richer in proteins related to cell division, microtubule organization, and RNA splicing; whereas in later stages, neuronal centrosome interactors are related to cytoskeleton and RNA-interacting proteins. Interestingly, the neural centrosome interactome is particularly enriched in RNA-interacting proteins compared with other cell types. By overlaying the interactomes with published datasets of neurodevelopmental diseases with de novo variants (DNV) in autism spectrum disorder (ASD), periventricular heterotopia (PH), intellectual disability (ID), epileptic encephalopathy (EE) and polymicrogyria (PMG), the authors detected a clear disease association of the neural centrosome interactome. In ASD, a pathological association was found for all datasets analysed, suggesting pan-cellular involvement of centrosome proteins in its aetiology. In PH, the authors identified the enrichment of the microtubule-anchoring pre-mRNA processing factor 6 (PRPF6). PRPF6 is more abundant in the centrosome of NSCs than of neurons. Mutated PRFF6 recapitulated PH heterotopias in the periventricular cortex of early mouse embryos, along altered mRNA splicing, that affected the centrosome associated Brsk2 (Brain-Selective Kinase 2) protein, involved in microtubule dynamic regulation and neural migration (Barnes et al., 2007; Kishi et al., 2005; Nakanishi et al., 2019). Indeed, RNA dynamics play an important function during brain development (Raj and Blencowe, 2015). Panagiotakos and Pasca, in a perspective manuscript in Neuron, remark how critical the moment and place of the events during brain development is for neurodevelopmental pathologies (Panagiotakos and Pasca, 2022). As an example, the temporal expression pattern of the voltage-gated sodium channels Nav1.1, Nav1.2, and Nav1.3 isoforms explains developmental brain malformations. Mutations in SCN3A, encoding for Nav1.3, which is elevated in immature progenitors and foetal brain neurons, can lead to abnormal neuronal migration and subsequent polymicrogyria (Smith et al., 2018). Instead, mutations in SCN1A and SCN2A encoding for Nav1.1 and Nav1.2, respectively (Beckh et al., 1989; Smith et al., 2018), which are elevated in postnatal neurons, are commonly related to infantile epilepsies (Meisler and Kearney, 2005). Interestingly, these protein isoforms also display specific cell-type enrichment during brain development. Parvalbumin (PV) cortical interneurons predominantly express Nav1.1 channels during early life (Yu et al., 2006), so that SCN1A loss of function leads to postnatal epilepsy, which disappears in adulthood (Favero et al., 2018). Thus, it is relevant to differentiate the initial mechanism that triggers disease onset, from those contributing to chronic disease states. In fact, the individual variability of neuropathology onset or affectation may depend on the moment or place the alteration occurs. Therefore, in addition to the proteomic and genetic information, understanding of neuropathology requires the understanding of cell-specific alterations, gene regulatory networks and protein interactomes and how they evolve and are regulated during the nervous system formation. 2. Stress granule assemblies and neurodevelopmental disorders Stress granules (SGs) are dynamic cytoplasmatic membrane-less compartments that assemble under a variety of stress conditions (Anderson and Kedersha, 2009; Jain et al., 2016). A large number of SGs components and regulators have been described (Jain et al., 2016; Markmiller et al., 2018; Yang et al., 2020), but the mechanistic dynamics of these assemblies are still unknown. Accumulated data indicates that these cytoplasmatic compartments play important roles in the regulation of gene expression (Buchan et al., 2008; Arimoto et al., 2008; Takahara and Maeda, 2012; Decker and Parker, 2012; Yang et al., 2020). SGs are detected where there are considerable pools of untranslated messengers and ribonucleoprotein particles (RNPs) (protein-coding mRNAs and non-protein-coding RNAs, and RNA-binding proteins) to shut down translation (Guillén-Boixet et al., 2020). Thus, they seem critical for gene expression homeostasis (Martin and Ephrussi, 2009; Wang et al., 2019), playing then relevant functions during brain development. Last year, Jia et al. reported a new NDD characterized by alterations in SG formation (Jia et al., 2022). They detected disruptive variants of UBAP2L, an essential regulator of SG formation (Youn et al., 2018; Cirillo et al., 2020), in patients with speech-language problems, ID, motor delay, seizure, and with less prevalence in patients with ADHD, ASD, repetitive and aggressive behaviour, and anxiety, but without a defined NDD. The patients also presented morphological features such as facial dysmorphisms, visual impairment, hypotonia and hand and foot abnormalities. Using skin fibroblast cell cultures from two patients, they showed reduced levels of UBAP2L and fewer SGs under stressful conditions. The authors validated these observations in a cell line (HeLa) knockout (KO) for UBAP2L. Transfection of these KO cells with the UBAP2L mutants detected in patients also led to a reduction in SGs formation. Further experiments in Ubap2l KO mice showed increased mortality in embryonic stages and reduced brain size compared with wild-type (WT) Ubap2l+/+, and heterozygous (Het) Ubap2l+/- littermates. KO mice showed anomalous neocortex lamination and reduction in neuronal progenitor proliferation possibly linked to altered SG dynamics during cortical neurogenesis. Moreover, Ubap2l+/- animals showed impaired social novelty ability, abnormal spatial working memory, and more anxiety-like behaviour. All these data prompted the authors to analyse the enrichment of SG genes from published datasets of proteomics and high-throughput genome-wide screenings in curated NDD gene datasets, including from SFARI (Simons Foundation Autism Research Initiative) and DDG2P database (Development Disorder Genotype - Phenotype Database). They detected significant enrichment of SG genes, particularly SG core genes and RNA-binding proteins. They also examined specific SG genes that could be related to NDD from previously reported de novo mutations (DNMs), detecting 3410 variants in the coding regions of 843 SG genes. The statistical analysis showed enrichment of SG genes that clustered according to their network function, STRING database for protein-protein interaction (PPI), with some of the enriched genes that had not been previously implicated in NDDs. Although previous works have evidenced that stress conditions during embryonic stages increase the risk of NDDs (Kinney et al., 2008; Babenko et al., 2015; Fitzgerald et al., 2020; Chui et al., 2020), this is the first study to identify alterations in SG assemblies as a common neuropathological feature of NDDs with no defined aetiology. 3. CLIP, a newly discovered interneuron progenitor, explains the divergent phenotype in tuberous sclerosis complex disorder Tuberous sclerosis complex (TSC) is a rare genetic condition that causes benign tumours in different parts of the body mainly the brain, kidneys, heart, skin, lungs and eyes. In the brain, TSC-associated lesions include subependymal tumours at the lateral ventricle and cortical dysplastic lesions, namely cortical tubers. Both aberrant structures contain, among other cell types, giant cells (GCs), which are the histopathological hallmark of the disease (Ruppe et al., 2014; Gelot and Represa, 2020; Henske et al., 2016). These cells feature a large and central nucleus with peripheral chromatin and a prominent nucleolus, and Nissl substance and neurofibrils in the cytoplasm (Mizuguchi, 2007). The abnormally large size of GCs strongly indicates dysregulation of cell size control in TSC (Mizuguchi, 2007). Patients often develop TSC-associated neuropsychiatric disorders (TAND) which include ID, attention deficit/hyperactivity disorder (ADHD), aggressiveness, difficulties with communication and social interaction (ASD), epilepsy, seizures and psychiatric conditions (Thiele, 2010). TSC is produced by the mutation of either TSC1 or TSC2. These two genes encode for proteins that inhibit mTOR (mammalian target of rapamycin) signalling, which is the major regulator of cell growth. Loss of regulation of this signalling pathway leads to abnormal cell development and differentiation. Experimental data suggest that TSC is produced by a heterozygous germline mutation followed by somatic loss of heterozygosity (LOH) in the other allele, due to loss-of-function mutations (Crino, 2013; Feliciano et al., 2011; Feliciano et al., 2012). However, patient tissue analyses show that LOH occurs only in tumours and not in dysplastic tubers (Henske et al., 1996; Chan et al., 2004; Qin et al., 2010). Moreover, mouse models with LOH in either TSC1 or TSC2 cannot recapitulate the full spectrum of brain aberrations observed in patients. Last year, Eichmüller et al., solved the discrepancies owing to the discovery of a new interneuron progenitor (Eichmüller et al., 2022). The authors found that cerebral organoids derived from patients with TSC2+/- reproduced both histopathological features using different culture conditions; that is, brain tumours when cultured in high-nutrient medium, and dysplastic cortical tubers when cultured in low-nutrient medium. The characterization of the cellular composition by single-cell RNA-sequencing (scRNA-seq), along with exhaustive histological validation, allowed the authors to identify a specific interneuron progenitor population that gives rise to both the tumours and the cortical tuber lesions. Comparisons with human foetal brain data revealed that this interneuron progenitor is first detected in the caudal ganglionic eminence (CGE) during late mid-gestation, with manifest expansion and migration during late gestation. Given their origin and embryonic stage, the authors called these interneuron progenitor cells CLIP, for “caudal late interneuron progenitor”. CLIP cells seem to be particularly sensitive to mTOR levels, being disturbed upon loss of one copy of TSC1/2, which resulted in the over-proliferation of these progenitor cells. The authors determined that the tubers are generated from migrated CLIP interneurons while the tumours grow in the CGE as a consequence of an additional aberration in the second allele, most probably produced by the over-proliferation of these CLIP cells and the contribution of other factors or cell types (Figure 2). Thus, although derived from the same altered progenitor CLIP, tuber cells do not show LOH as a mechanism of action, while the tumour cells do show LOH.
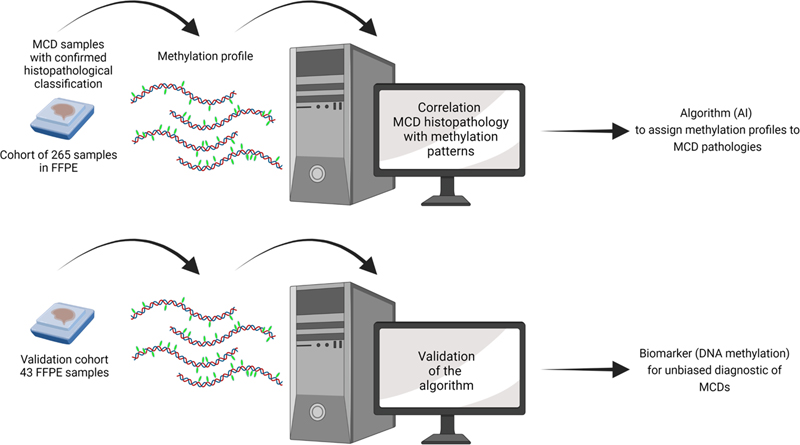
Figure 2. Illustration depicting the mechanism described by Eichmüller et al., 2022. The left shows normal development when neither of the two copies of TSC1/2 have mutations. On the right, when one copy of TSC1/2 is mutated, CLIP cells become sensitive to mTOR levels, resulting in aberrant growth and expansion. CLIP neurons that migrate to the cortex develop into cortical tubers, and the remaining CLIP cells, through the participation of other additional alterations lose the other allele producing tumours. The manuscript shows how the same progenitor cell type diverges into two histopathological differential phenotypes. It also shows that CLIP cells depend on epidermal growth factor receptor (EGFR) signalling, and that the inhibition of EGFR regressed the organoid tumours, providing an alternative treatment therapy for this pathology. An interesting aspect of this manuscript is that the disease mechanism described is human-specific. Indeed, human brain development encompasses the generation and/or expansion of cell types deriving large and gyrated cortices, which do not occur in small lissencephalic brains such as the rodent brain cortex. Even postnatally there is extensive migration of interneurons from the CGE into the cortex in humans (Paredes et al., 2016; Hansen et al., 2013; Hodge et al., 2019), but not in mice (Raju et al., 2018). The use of human organoids was key for this discovery. However, although human organoids are a powerful model system, this technology is still in its infancy. For example, the current lack of standardized protocols implies important variability among organoids from the same patient, which intrinsically puts the results in uncertainty. Thus, further studies are necessary to validate CLIP cells and their functions. 4. Improving the diagnostic of malformations of cortical development (MCDs) diseases by DNA methylation patterns Malformations of cortical development (MCDs) comprise various neurodevelopmental disorders that are a major cause of epilepsy (Leventer et al., 1999), and medically stubborn childhood seizures (Kuzniecky, 1995). MCDs can be classified into three groups depending on their likely origin. In group I, derived from abnormal cell proliferation or apoptosis, there are hemi-megalencephaly, microcephaly, megalencephaly, and focal cortical dysplasia; in group II, related to abnormal cell migration, we find tubulinopathies, lissencephalies and heterotopies; and in group III polymicrogyria is produced by abnormal post-migrational development (Desikan and Barkovich, 2016). This heterogeneity of causes and phenotypic presentations with a broad range of symptomatology including cognitive deficits, ID, and ASD (Barkovich et al., 2012; Guerrini and Dobyns, 2014), challenges neuropathologists in providing an accurate diagnosis and, consequently, an on-target prognosis and management of the affectation. As such, biomarkers to identify the type of MCD more precisely are a growing subject of research. However, biomarkers are available only for focal cortical dysplasia (FCD) type II (D’Gama et al., 2015; Jansen et al., 2015; D’Gama et al., 2017; Baldassari et al., 2019) and mild malformations of cortical development with oligodendroglial hyperplasia (MOGHE) (Schurr et al., 2017; Bonduelle et al., 2021). Currently, the diagnosis of MCD pathologies is based only on histopathological criteria providing, generally, an imprecise diagnosis. As an example, in the case of FCD type II, two forms have been described - FCDIIA and FCDIIB - (Blümcke et al., 2011), which differ in that only FCDIIB contains balloon cells (BCs). BCs are enlarged cytoplasm cells that resemble gemistocytic astrocytes displaying multiple or convoluted nuclei without prominent nucleoli (Mizuguchi, 2007). However, BCs histologically are very alike to GCs observed in TSC. FCDIIB and TSC are both pathologies associated with dysregulation of the mTOR pathway and display comparable histopathological features, particularly FCDIIB and the cortical tubers, which suggests a closely related origin, although they are clearly different neuropathological entities (Taylor et al., 1971; Lee et al., 2022). In an attempt to improve the diagnosis of MCD pathologies, Jabari et al. assayed a potential strategy based on DNA methylation (Jabari et al., 2022). DNA methylation can be a reliable biomarker because it is preserved and, therefore, can be detected in archival human brains stored in FFPE (Sahm et al., 2017; Capper et al., 2018; Wefers et al., 2020). Moreover, the methylome manifests a combination of both the somatically acquired DNA methylation alterations, and the molecular memory marks in response to environmental or pathogenic cues (Kobow and Blümcke, 2012; Kobow et al., 2013; Kiese et al., 2017; Kobow et al., 2019; Kobow et al., 2020). Furthermore, DNA methylation profile is widely used to classify CNS tumours because of its reproducibility and sensitivity even in small samples (Sahm et al., 2017; Capper et al., 2018). Thus, the purpose of this study was to find DNA methylation patterns to accurately classify the different histopathological entities. The authors used surgical samples from patients with MCD and with a confirmed histopathological classification included 265 samples across all age groups and sex that demonstrated different pathological levels of the 12 major subtypes of MCD along with different controls. The authors performed a genome-wide DNA methylation assay to correlate the DNA methylation patterns with the histopathological classification. They used three different approaches: pairwise comparison, machine learning, and deep learning algorithms. The deep learning algorithm allowed for the most accurate discrimination providing a rationalized classification of the pathologies. Then, they analysed the precision of the DNA methylation-based MCD classification using a new cohort from different epilepsy centres. This test cohort contained 43 surgical FFPE samples, among which some previously underwent multiple rounds of histopathological evaluation from expert neuropathologists because of the difficulty of their classification. Using the algorithm, the authors were able to accurately classify all samples from the test cohort. Figure 3 depicts the flowchart the authors followed. Thus, they demonstrate that DNA methylation-based MCD classification is suitable across major histopathological entities and could be used to establish an integrated diagnostic classification scheme for MCD neuropathology.
Figure 3. Flowchart of the process. Top, generation of the algorithm by artificial intelligence (AI) of the correlation of the DNA methylation profiles obtained from FFPE samples with confirmed histopathological MCDs classification. Bottom, validation of the algorithm to classify MCDs based on DNA methylation using a new set of FFPE samples. 5. Deciphering the aetiology of sudden unexplained death in childhood (SUDC) by proteomics Sudden unexplained death in childhood (SUDC) refers to the unexplainable death of children over 1 year of age. It is called unexplained because, after a complete review of the clinical history and the autopsy including toxicologic, genetic, metabolic and microbiology analyses, to cite some of the complementary studies, the cause of death is not determined. Although the causes may be diverse, genetic variants are likely to be prone to SUDC risk (Crandal et al., 2020; Halvorsen et al., 2021; Harowitz et al., 2021; Holm et al., 2012; Narula et al., 2015). For example, among SUDC cases, there is a high prevalence of individual or familial febrile seizure (FS) history (Hefti et al., 2016; Hesdorffer et al., 2015; McGuone et al., 2020). SUDC shares some pathological similarities with sudden unexpected death in epilepsy (SUDEP) (Devinsky et al., 2016; Kinney et al., 2016), suggesting that they may share some coincident mechanisms that result in premature death. Similar to SUDC, the causes of SUDEP may also be diverse including genetic risk factors. Since both pathological conditions show abnormalities in the hippocampus and cortex (Ackerman et al., 2016; Kinney et al., 2016; Kon et al., 2020; McGuone et al., 2020), these brain regions, particularly the hippocampus, have attracted most SUDC studies. However, no conclusive results are available to date (Leitner et al., 2022a; Roy et al., 2020). In this scenario, Leitner et al., have defined differential protein abundance in several brain areas of SUDC cases (19 cases), including the frontal cortex, hippocampal dentate gyrus (DG), and cornu ammonis (CA1-3). The study compared cases with and without febrile seizure history (SUDC-FS and SUDC-noFS) and without febrile seizure with control cases (n = 19) matched by age, sex, brain weight and post-mortem interval (Leitner et al., 2022b). The authors micro-dissected the aforementioned brain regions from autopsy FFPE tissue to perform label-free quantitative proteomic analyses. The proteomic analyses revealed no differential hippocampal neuropathology between SUDC-FC and SUDC-noFC. Instead, principal component analysis (PCA) revealed a significant separation between SUDC and controls in the frontal cortex, but not regarding FS history. Differential protein abundance analysis showed significant differences between SUDC and control cases in 660 proteins of the frontal cortex, while only 170 in the DG and 57 in CA1-3. Pathway analysis revealed 238 signalling pathways in the frontal cortex, mainly involved in the activation of oxidative phosphorylation, inhibition of EIF2 (Eukaryotic Initiation Factor 2) signalling, and glutamate receptor signalling. In the DG, they mainly found pathways involving activation of the acute phase response and inhibition of reelin signalling, while in hippocampal CA1-3, the only involved signalling pathway was in the acute phase response associated with cellular stress response (Liu and Qian, 2014; Brace et al., 2016; Leitner et al., 2022b). The fact that they detected protein overlap in more than one signalling pathway supports the involvement of these signalling pathways. The authors also used weighted gene correlation network analysis (WGCNA) to correlate the proteomic results with the clinical history. In the frontal cortex there were common affected enriched signaling pathways in SUDC and SUDEP, some of which showed oppository effects in SUDC and SUDEP (e.g., oxidative phosphorylation and EIF2 signalling), while other proteomic pathways were similar(e.g., mitochondrial enzyme COX6B1). As a corollary, besides the relevant information provided, one of the most remarkable findings is that while the frontal cortex is not a studied region in SUDC, it showed the most altered proteomic changes. Hence, the authors put forward the relevance of the frontal cortex in this condition and the necessity of performing studies in this region to detect possible neuropathological signs associated with SUDC to reduce and prevent the risk of this condition. 6. Transcriptomic dysregulation in ASD occurs across the whole cerebral cortex and follows a regional gradient ASD is a prototypical example of neuropathology in which a definitive aetiology has not yet been determined. In the last years, comprehensive omics assessments have been conducted to determine risk genes (de la Torre-Ubieta et al., 2016) or differential patterns of splicing or gene isoform expression in ASD (Wu et al., 2016; Sun et al., 2016; Gandal et al., 2018). Despite the heterogeneity of factors that drive to ASD pathology, molecular profiling studies have found consistent patterns of transcriptomic and epigenetic dysregulation (Ramaswami et al., 2020), involving upregulation of astrocytes, microglia, and neural immune genes; and downregulation of synaptic, neurite morphogenesis, and neuronal energy pathway genes accompanied by attenuation of gene-expression gradients in cortical association regions (Voineagu et al., 2011; Wu et al., 2016; Parikshak et al., 2016; Gandal et al., 2018). The work of Gandal et al. explores whether these alterations are more widespread throughout the cortex and proposes that ASD pathology is the physiological manifestation of altered cortical arealization (Gandal et al., 2022). To support this hypothesis, they did bulk RNA-sequencing (RNA-seq) analysis to identify altered genes and alternative spliced gene isoforms. They used threefold more samples than previous works (Voineagu et al., 2011; Parikshak et al., 2016), analysing 725 post-mortem brain samples spanning 11 cortical areas from 112 individuals of both sexes and with ages ranging from 2 to 68, totalling 49 subjects with idiopathic ASD and 54 neurotypical controls. They found transcriptomic changes across the cortex of ASD patients with an anterior-to-posterior gradient, with the most remarkable differences in the primary visual cortex. In agreement with previous reports (Walker et al., 2019), the greater differential expression detected was related to alternative splicing and differential isoform expression. Thus, their findings indicate that molecular alterations, mainly in alternative splicing and isoform expression, in ASD transcriptome extend beyond the associative cortex to broadly involve primary sensory regions. This may explain the altered sensory processing observed in individuals with ASD. An interesting aspect is that the differences in gene expression that account for demarcating the different cortical regions, because they define the cytoarchitecture, connectivity, and function of a particular region, were attenuated in ASD. This indicates that the cortical regions in ASD patients are molecularly more homogeneous, and therefore, less differentiated and specialised. Again, this attenuated expression followed a gradient pattern, which was more particularly affected in the posterior regions such as the primary visual cortex. The authors also determined whether the transcriptomic changes detected were reflected in the cell-specific type gene expression. By sn-RNA-seq and methylation profiles, they determined a substantial differential expression profile in excitatory neuron classes and glia cells, once more, with a regional gradient more prominent in occipital and parietal cell types than in prefrontal cortex (PFC). This reiterative regional gradient may reflect a reminiscence of the buildout of the cortical cytoarchitecture, i.e., its patterning and connectivity, which depend on both cell intrinsic factors genetic and epigenetic regulatory programmes, and extrinsic signals, –such as morphogen gradients (Cadwell et al., 2019). The data suggests that this process is altered in ASD, indicating an early developmental alteration in cortical arealization, affecting local neuronal circuits, synaptic homeostasis, and leading to the ASD manifestations. 7. In vivo platform for understanding the neuropathology of human neurodevelopmental diseases Some years ago, several studies showed that it was possible to transplant human neurons into the rodent cortex that were able to establish connections with the rodent cells (Espuny-Camacho et al., 2013; Mansour et al., 2018; Real et al., 2018; Linaro et al., 2019; Kitahara et al., 2020; Xiong et al., 2021), thus providing an in vivo platform to study human developmental disorders. Although the hope was that those tools would allow uncovering circuit-level phenotypes from patient-derived cells and test therapeutic strategies, they had some problems. Part of those have now been solved in the work of Revah et al. (Revah et al., 2022). To facilitate integration of the transplant they transplanted 3D human cortical organoids (hCO) into the somatosensory cortex of immunodeficient rats, at early postnatal stage, in which corticocortical and thalamocortical innervation have not yet been completed, thus minimising the endogenous circuitry alteration (Kichula and Huntley, 2008). The novelty of this work is thus that they transplanted intact organoids, rather than a dissociated cell suspension; and at very early postnatal stages, rather than in adult rats. This strategy allows better synaptic and axonal integration of the human cells. Hence, the transplanted hCOs (t-hCO) displayed more mature properties compared to not transplanted same-age hCOs. The t-hCO showed vascularization and the presence of microglia. SnRNA-seq revealed the canonical expression pattern of major cortical cell classes, even though t-hCO did not present anatomical lamination. Transplanted neurons showed a more mature morphological phenotype, i.e., larger somas, more dendrites, larger processes, and higher dendritic spine density, and more mature electrophysiological properties. The authors traced t-hCO innervation with retrograde rabies showing integration of t-hCO neurons in the rat brain's circuitry. Fibre photometry, two-photon calcium imaging, electrophysiology, and sensory stimuli by deflecting the rat’s whiskers revealed the functionality of the integration, as t-hCO stimulation evoked response in rat neurons indicating functional innervation. Finally, by in vivo optogenetics they showed that activation of t-hCO could modify the rats’ behavioural response. The authors used this system for studying Timothy Syndrome (TS), a severe genetic disorder caused by the mutation in the L-type voltage-sensitive calcium channel CaV1.2 (Ebert and Greenberg, 2013). They compared the evolution of t-hCO derived from control individuals and patients with TS with their non-transplanted counterparts (hCO). TS t-hCO neurons showed altered dendritic morphology with an extensive number of primary dendrites but with an overall reduction in the mean and total dendritic size. Also, these neurons displayed increased synaptic spine density that impaired their electrophysiological properties. These phenotypes could only be detected in TS t-hCO but not in non-transplanted organoids with the same differentiation stage, thus indicating that organoid transplantation allowed for better recapitulation of the disease phenotype that was elusive in non-transplanted organoids. 8. Mitochondria participate in the neuron-glia crosstalk Mitochondria play crucial roles in the regulation of cellular energy and metabolism; therefore, it is not surprising that this organelle is involved in all developmental stages with important functions in neuronal differentiation. For this reason, several mitochondrial disorders present an abnormal neuronal and neurological development (Son and Han, 2018). Mitochondrial dysfunction has been widely associated with neurological and psychiatric diseases such as schizophrenia, bipolar depression, ASD and Rett syndrome (Son and Han, 2018). One example is Leigh syndrome, which is caused by a mutation in mitochondrial DNA, leading to dysfunctional mitochondrial complexes. The disorder usually manifests within the first year of life and leads to rapid degeneration of physical and mental abilities, ultimately leading to death within 2-3 years (Murphy and Craig, 1975). However, this close relationship between the mitochondria and the CNS is not only applicable to neurons. In fact, it is astrocytic’ mitochondria that support neuronal development and function. In cases of neuronal damage, such as in brain ischemia, astrocytes release functional mitochondria into the extracellular medium via a mechanism that involves the activation of the cluster of differentiation 38 (CD38) and cyclic adenosine diphosphate (ADP)-ribose signalling (Hayakawa et al., 2016; English et al., 2020). This process facilitates neuronal recovery. At the same time, damaged neurons expel damaged mitochondria, which are in turn absorbed by the surrounding astrocytes to be recycled. In a paper published last year, Gao et al. showed that the release of mitochondria might function as a signalling element between neurons and glial cells (Gao et al., 2022). They also proved that, under physiological conditions, neurons release mitochondria into the extracellular medium. Then, they tested different pathological conditions in vitro, such as acidosis, hydrogen peroxide or high levels of NMDA (N-methyl D-aspartate) or glutamate, simulating brain ischemia. They detected a significant increase in the number of released damaged mitochondria by the neurons (Gao et al., 2022), and proved that astrocytes uptake these mitochondria and trigger the rescuing response. Hence, they speculate that, under stress conditions, mitochondria could serve as a “help-me” signal. If replicated, this discovery would represent a huge step forward in understanding the mechanisms behind neuronal stress response that could shed light on common neurodevelopmental/neurodegenerative disorders. 9. Gaining insight on the role of HIV-1 in the CNS Since its discovery, the human immunodeficiency virus (HIV) has been associated with several neurological conditions. HIV-1 infection has long been known to have an impact on the nervous system, culminating in HIV-associated neurocognitive disorder (HAND) (Clifford and Ances, 2013), as the brain, along with the bone marrow, is known to be a reservoir for the virus. An interesting aspect is how HIV-1 infection affects neurodevelopment. In the era of antiretroviral therapy (ART), this question translates to how maternal infection affects offspring development. Properly treated mothers give birth to uninfected children in almost 99% of cases (Wedderburn et al., 2022). However, HIV-exposed children generally show a slower development of some neurological functions. In particular, a poorer expressive language and gross motor function compared to their HIV-unexposed counterparts (Wedderburn et al., 2022). However, the mechanisms underlying this phaenomenon remain unclear. One hypothesis states that the virus could directly affect the development of the foetus, while other researchers think that it is the chronic systemic inflammation of the mother due to the infection what could affect the offspring. However, research has been hindered by the lack of a satisfactory model, owing to the complexity derived from the involvement of both the nervous and immune systems. For this reason, researchers have long been searching for a novel model to study this phenomenon. Brain organoids, in fact, could provide an insight on neuronal development and in recent years, protocols for microglia-containing brain organoids have been developed, hence allowing for this technology to be applied to HIV research field (Ormel et al., 2018). To prove so, a study in 2022 by Gumbs et al. showed that the cluster of differentiation 4 (CD4) and Cysteine-Cysteine chemokine receptor 5 (CCR5)– expressing microglia were susceptible to HIV-1 infection in a human brain organoid model (Gumbs et al., 2022). They tested this infection model on both microglia-containing organoids and organoid-derived microglia (oMG), with positive results in both cases. Albeit the obvious limitations of the organoid model, the results of this study represent a significant advancement in the HIV research field, as these organoids could help elucidate the mechanisms behind the effects of the virus on the brain and test novel therapies. 10. Novel mechanisms explaining COVID-19 neurological anomalies The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) pandemic has had a huge impact on everyone's lives, causing millions of deaths worldwide and changing perpetually our society. Although the disease was initially known to be hazardous to the respiratory system, soon several comorbidities with long-lasting effects were associated with the virus. It was soon discovered that the infection could lead to several significant neurological complications (Chen et al., 2022). These effects vary largely from patient to patient, depending on factors such as age, weight, health status and pre-existing conditions. However, both short- and long-lasting neurological effects have been reported in all populations, from children to adults. Regarding the effects of COVID-19 on children, we should distinguish between the effects due to the pandemic and those due to the direct effects of the virus. A meta-analysis study by Hessami et al. highlighted that infants born or raised during the pandemic showed a higher rate of communication impairment than the pre-pandemic cohort (Hessami et al., 2022). Given the nature of the study and the lack of other significant neurodevelopmental impairments in the general paediatric population, we can speculate that this communication impairment was an effect of the lack of social stimuli because of the lockdown rather than the virus itself. However, the direct neurological consequences of COVID-19 infection have also been reported in both children and adults. These effects range from temporary anosmia and ageusia to memory loss, meningitis, stroke, and neurodegeneration (Ledford, 2022). Data on the possible long-term neurological effects of SARS-CoV-2 coronavirus are inconclusive. Several studies have found associations with Alzheimer’s disease (AD) risk, cardiovascular damage, or neurological sequelae. An analysis in the UK found a slight overall reduction of grey matter in the brains of recovered individuals. Also, that the virus infects brain support cells and induces inflammation similar to Parkinson’s or AD, and that severe COVID-19 causes detectable brain ageing (Douaud et al., 2022). Despite the significant amount of research conducted on and the neurotropic nature of the virus, the molecular mechanisms linking COVID-19 to these neurological symptoms are still unclear. A study published in 2022 by Charnley et al. showed that some viral open reading frame (ORF) proteins can assemble into amyloid-like aggregates and cause neurotoxicity (Charnley et al., 2022). Using an algorithm, they pinpointed two short regions of ORF6 and ORF10 as those responsible for the assembly. They then tested the peptides corresponding to these regions and showed that they self-assembles into amyloid-like structures. Finally, these peptides were tested on neuroblastoma SH-SY5Y cells, proving to be neurotoxic (Charnley et al., 2022). We can presume that some of the neurological consequences of a COVID-19 infection not only share symptomatology with some common neurodegenerative disorders (e.g., AD), but also share an accumulated misfolding protein-driven pathogenesis. It will be now important to test these mechanisms during development. Funding The lab of M.D. is supported by the Departament d'Universitats, Recerca i Societat de la Informació de la Generalitat de Catalunya (Grups consolidats 2022), the Agencia Estatal de Investigación (PID2019-110755RB-I00/AEI/10.13039/501100011033), the European Union’s Horizon 2020 research and innovation programme under grant agreements nº 848077 (GO-DS21) and under grant agreement nº 899986 (ICOD). Horizon Europe (Horizon-Hlth-2021-StayHlth-01-Psych-STRATA). This reflects only the author's view, and the European Commission is not responsible for any use that may be made of the information it contains. Jerôme Lejeune Foundation (Grant number 2002), NIH (Grant Number: 1R01EB 028159-01), Marató TV3 (#2022-12-30-31-32). The CRG acknowledges the support of the Spanish Ministry of Science and Innovation to the EMBL partnership, the Centro de Excelencia Severo Ochoa, and the CERCA Programme/Generalitat de Catalunya. The CIBER of Rare Diseases (CIBERER) is an initiative of ISCIII. References Ackerman MJ, Andrew TA, Baker AM, Devinsky O, Downs JC, Keens T, Kuntz J, Lin P, Lear-Kaul KC, Reichard R, Robinson DA. An Association of Hippocampal Malformations and Sudden Death? We Need More Data. Forensic Sci Med Pathol. 2016 Jun;12(2):229-31. https://doi.org/10.1007/s12024-016-9765-1. Epub 2016 Mar 26. PMID: 27017493. Anderson P, Kedersha N. RNA granules: post-transcriptional and epigenetic modulators of gene expression. Nat Rev Mol Cell Biol. 2009 Jun;10(6):430-6. https://doi.org/10.1038/nrm2694. PMID: 19461665. Arimoto K, Fukuda H, Imajoh-Ohmi S, Saito H, Takekawa M. Formation of stress granules inhibits apoptosis by suppressing stress-responsive MAPK pathways. Nat Cell Biol. 2008 Nov;10(11):1324-32. https://doi.org/10.1038/ncb1791. Epub 2008 Oct 5. PMID: 18836437. Babenko O, Kovalchuk I, Metz GA. Stress-induced perinatal and transgenerational epigenetic programming of brain development and mental health. Neurosci Biobehav Rev. 2015 Jan;48:70-91. https://doi.org/10.1016/j.neubiorev.2014.11.013. Epub 2014 Nov 24. PMID: 25464029. Baldassari S, Ribierre T, Marsan E, Adle-Biassette H, Ferrand-Sorbets S, et al. Dissecting the genetic basis of focal cortical dysplasia: a large cohort study. Acta Neuropathol. 2019 Dec;138(6):885-900. https://doi.org/10.1007/s00401-019-02061-5. Epub 2019 Aug 23. PMID: 31444548; PMCID: PMC6851393. Barkovich AJ, Guerrini R, Kuzniecky RI, Jackson GD, Dobyns WB. A developmental and genetic classification for malformations of cortical development: update 2012. Brain. 2012 May;135(Pt 5):1348-69. https://doi.org/10.1093/brain/aws019. Epub 2012 Mar 16. PMID: 22427329; PMCID: PMC3338922. Barnes AP, Lilley BN, Pan YA, Plummer LJ, Powell AW, Raines AN, Sanes JR, Polleux F. LKB1 and SAD kinases define a pathway required for the polarization of cortical neurons. Cell. 2007 May 4;129(3):549-63. https://doi.org/10.1016/j.cell.2007.03.025. PMID: 17482548. Beckh S, Noda M, Lübbert H, Numa S. Differential regulation of three sodium channel messenger RNAs in the rat central nervous system during development. EMBO J. 1989 Dec 1;8(12):3611-6. https://doi.org/10.1002/j.1460-2075.1989.tb08534.x. PMID: 2555170; PMCID: PMC402042. Blümcke I, Thom M, Aronica E, Armstrong DD, Vinters HV, et al. The clinicopathologic spectrum of focal cortical dysplasias: a consensus classification proposed by an ad hoc Task Force of the ILAE Diagnostic Methods Commission. Epilepsia. 2011 Jan;52(1):158-74. https://doi.org/10.1111/j.1528-1167.2010.02777.x. Epub 2010 Nov 10. PMID: 21219302; PMCID: PMC3058866. Bonduelle T, Hartlieb T, Baldassari S, Sim NS, Kim SH, et al. Frequent SLC35A2 brain mosaicism in mild malformation of cortical development with oligodendroglial hyperplasia in epilepsy (MOGHE). Acta Neuropathol Commun. 2021 Jan 6;9(1):3. https://doi.org/10.1186/s40478-020-01085-3. PMID: 33407896; PMCID: PMC7788938. Bonnefont J, Vanderhaeghen P. Neuronal fate acquisition and specification: time for a change. Curr Opin Neurobiol. 2021 Feb;66:195-204. https://doi.org/10.1016/j.conb.2020.12.006. Epub 2021 Jan 5. PMID: 33412482; PMCID: PMC8064025. Brace LE, Vose SC, Stanya K, Gathungu RM, Marur VR, et al. Increased oxidative phosphorylation in response to acute and chronic DNA damage. NPJ Aging Mech Dis. 2016 Oct 13;2:16022. https://doi.org/10.1038/npjamd.2016.22. PMID: 28721274; PMCID: PMC5514997. Buchan JR, Muhlrad D, Parker R. P bodies promote stress granule assembly in Saccharomyces cerevisiae. J Cell Biol. 2008 Nov 3;183(3):441-55. https://doi.org/10.1083/jcb.200807043. PMID: 18981231; PMCID: PMC2575786. Cadwell CR, Bhaduri A, Mostajo-Radji MA, Keefe MG, Nowakowski TJ. Development and Arealization of the Cerebral Cortex. Neuron. 2019 Sep 25;103(6):980-1004. https://doi.org/10.1016/j.neuron.2019.07.009. PMID: 31557462; PMCID: PMC9245854. Capper D, Jones DTW, Sill M, Hovestadt V, Schrimpf D, et al. DNA methylation-based classification of central nervous system tumours. Nature. 2018 Mar 22;555(7697):469-474. https://doi.org/10.1038/nature26000. Epub 2018 Mar 14. PMID: 29539639; PMCID: PMC6093218. Chan JA, Zhang H, Roberts PS, Jozwiak S, Wieslawa G, Lewin-Kowalik J, Kotulska K, Kwiatkowski DJ. Pathogenesis of tuberous sclerosis subependymal giant cell astrocytomas: biallelic inactivation of TSC1 or TSC2 leads to mTOR activation. J Neuropathol Exp Neurol. 2004 Dec;63(12):1236-42. https://doi.org/10.1093/jnen/63.12.1236. PMID: 15624760. Charnley M, Islam S, Bindra GK, Engwirda J, Ratcliffe J, Zhou J, Mezzenga R, Hulett MD, Han K, Berryman JT, Reynolds NP. Neurotoxic amyloidogenic peptides in the proteome of SARS-COV2: potential implications for neurological symptoms in COVID-19. Nat Commun. 2022 Jun 13;13(1):3387. https://doi.org/10.1038/s41467-022-30932-1. PMID: 35697699; PMCID: PMC9189797. Chen Y, Yang W, Chen F, Cui L. COVID-19 and cognitive impairment: neuroinvasive and blood‒brain barrier dysfunction. J Neuroinflammation. 2022 Sep 7;19(1):222. https://doi.org/10.1186/s12974-022-02579-8. PMID: 36071466; PMCID: PMC9450840. Chui A, Zhang Q, Dai Q, Shi SH. Oxidative stress regulates progenitor behavior and cortical neurogenesis. Development. 2020 Mar 11;147(5):dev184150. https://doi.org/10.1242/dev.184150. PMID: 32041791; PMCID: PMC7075051. Cirillo L, Cieren A, Barbieri S, Khong A, Schwager F, Parker R, Gotta M. UBAP2L Forms Distinct Cores that Act in Nucleating Stress Granules Upstream of G3BP1. Curr Biol. 2020 Feb 24;30(4):698-707.e6. https://doi.org/10.1016/j.cub.2019.12.020. Epub 2020 Jan 16. PMID: 31956030. Clifford DB, Ances BM. HIV-associated neurocognitive disorder. Lancet Infect Dis. 2013 Nov;13(11):976-86. https://doi.org/10.1016/S1473-3099(13)70269-X. PMID: 24156898; PMCID: PMC4108270. Crandall LG, Lee JH, Friedman D, Lear K, Maloney K, et al. Evaluation of Concordance Between Original Death Certifications and an Expert Panel Process in the Determination of Sudden Unexplained Death in Childhood. JAMA Netw Open. 2020 Oct 1;3(10):e2023262. https://doi.org/10.1001/jamanetworkopen.2020.23262. PMID: 33125496; PMCID: PMC7599447. Crino PB. Evolving neurobiology of tuberous sclerosis complex. Acta Neuropathol. 2013 Mar;125(3):317-32. https://doi.org/10.1007/s00401-013-1085-x. Epub 2013 Feb 6. PMID: 23386324. de la Torre-Ubieta L, Won H, Stein JL, Geschwind DH. Advancing the understanding of autism disease mechanisms through genetics. Nat Med. 2016 Apr;22(4):345-61. https://doi.org/10.1038/nm.4071. PMID: 27050589; PMCID: PMC5072455. Decker CJ, Parker R. P-bodies and stress granules: possible roles in the control of translation and mRNA degradation. Cold Spring Harb Perspect Biol. 2012 Sep 1;4(9):a012286. https://doi.org/10.1101/cshperspect.a012286. PMID: 22763747; PMCID: PMC3428773. Delgehyr N, Sillibourne J, Bornens M. Microtubule nucleation and anchoring at the centrosome are independent processes linked by ninein function. J Cell Sci. 2005 Apr 15;118(Pt 8):1565-75. https://doi.org/10.1242/jcs.02302. Epub 2005 Mar 22. PMID: 15784680. Desikan RS, Barkovich AJ. Malformations of cortical development. Ann Neurol. 2016 Dec;80(6):797-810. https://doi.org/10.1002/ana.24793. Epub 2016 Nov 11. PMID: 27862206; PMCID: PMC5177533. Devinsky O, Hesdorffer DC, Thurman DJ, Lhatoo S, Richerson G. Sudden unexpected death in epilepsy: epidemiology, mechanisms, and prevention. Lancet Neurol. 2016 Sep;15(10):1075-88. https://doi.org/10.1016/S1474-4422(16)30158-2. Epub 2016 Aug 8. PMID: 27571159. D'Gama AM, Geng Y, Couto JA, Martin B, Boyle EA, et al. Mammalian target of rapamycin pathway mutations cause hemimegalencephaly and focal cortical dysplasia. Ann Neurol. 2015 Apr;77(4):720-5. https://doi.org/10.1002/ana.24357. Epub 2015 Feb 26. PMID: 25599672; PMCID: PMC4471336. D'Gama AM, Woodworth MB, Hossain AA, Bizzotto S, Hatem NE, et al. Somatic Mutations Activating the mTOR Pathway in Dorsal Telencephalic Progenitors Cause a Continuum of Cortical Dysplasias. Cell Rep. 2017 Dec 26;21(13):3754-3766. https://doi.org/10.1016/j.celrep.2017.11.106. PMID: 29281825; PMCID: PMC5752134. Di Bella DJ, Habibi E, Stickels RR, Scalia G, Brown J, et al. Molecular logic of cellular diversification in the mouse cerebral cortex. Nature. 2021 Jul;595(7868):554-559. https://doi.org/10.1038/s41586-021-03670-5. Epub 2021 Jun 23. Erratum in: Nature. 2021 Aug;596(7873):E11. PMID: 34163074; PMCID: PMC9006333. Douaud G, Lee S, Alfaro-Almagro F, Arthofer C, Wang C, McCarthy P, Lange F, Andersson JLR, Griffanti L, Duff E, Jbabdi S, Taschler B, Keating P, Winkler AM, Collins R, Matthews PM, Allen N, Miller KL, Nichols TE, Smith SM. SARS-CoV-2 is associated with changes in brain structure in UK Biobank. Nature. 2022 Apr;604(7907):697-707. https://doi.org/10.1038/s41586-022-04569-5. Epub 2022 Mar 7. PMID: 35255491; PMCID: PMC9046077. Ebert DH, Greenberg ME. Activity-dependent neuronal signalling and autism spectrum disorder. Nature. 2013 Jan 17;493(7432):327-37. https://doi.org/10.1038/nature11860. PMID: 23325215; PMCID: PMC3576027. Eichmüller OL, Corsini NS, Vértesy Á, Morassut I, Scholl T, et al. Amplification of human interneuron progenitors promotes brain tumors and neurological defects. Science. 2022 Jan 28;375(6579):eabf5546. https://doi.org/10.1126/science.abf5546. Epub 2022 Jan 28. PMID: 35084981; PMCID: PMC7613689. English K, Shepherd A, Uzor NE, Trinh R, Kavelaars A, Heijnen CJ. Astrocytes rescue neuronal health after cisplatin treatment through mitochondrial transfer. Acta Neuropathol Commun. 2020 Mar 20;8(1):36. https://doi.org/10.1186/s40478-020-00897-7. PMID: 32197663; PMCID: PMC7082981. Espuny-Camacho I, Michelsen KA, Gall D, Linaro D, Hasche A, et al. Pyramidal neurons derived from human pluripotent stem cells integrate efficiently into mouse brain circuits in vivo. Neuron. 2013 Feb 6;77(3):440-56. https://doi.org/10.1016/j.neuron.2012.12.011 . PMID: 23395372. Favero M, Sotuyo NP, Lopez E, Kearney JA, Goldberg EM. A Transient Developmental Window of Fast-Spiking Interneuron Dysfunction in a Mouse Model of Dravet Syndrome. J Neurosci. 2018 Sep 5;38(36):7912-7927. https://doi.org/10.1523/JNEUROSCI.0193-18.2018. Epub 2018 Aug 13. PMID: 30104343; PMCID: PMC6125809. Feliciano DM, Quon JL, Su T, Taylor MM, Bordey A. Postnatal neurogenesis generates heterotopias, olfactory micronodules and cortical infiltration following single-cell Tsc1 deletion. Hum Mol Genet. 2012 Feb 15;21(4):799-810. https://doi.org/10.1093/hmg/ddr511. Epub 2011 Nov 7. PMID: 22068588; PMCID: PMC3263992. Feliciano DM, Su T, Lopez J, Platel JC, Bordey A. Single-cell Tsc1 knockout during corticogenesis generates tuber-like lesions and reduces seizure threshold in mice. J Clin Invest. 2011 Apr;121(4):1596-607. https://doi.org/10.1172/JCI44909. Epub 2011 Mar 14. PMID: 21403402; PMCID: PMC3069783. Fitzgerald E, Hor K, Drake AJ. Maternal influences on fetal brain development: The role of nutrition, infection and stress, and the potential for intergenerational consequences. Early Hum Dev. 2020 Nov;150:105190. https://doi.org/10.1016/j.earlhumdev.2020.105190. Epub 2020 Sep 10. PMID: 32948364; PMCID: PMC7481314. Gandal MJ, Haney JR, Wamsley B, Yap CX, Parhami S, et al. Broad transcriptomic dysregulation occurs across the cerebral cortex in ASD. Nature. 2022 Nov;611(7936):532-539. https://doi.org/10.1038/s41586-022-05377-7. Epub 2022 Nov 2. PMID: 36323788; PMCID: PMC9668748. Gandal MJ, Zhang P, Hadjimichael E, Walker RL, Chen C, et al. Transcriptome-wide isoform-level dysregulation in ASD, schizophrenia, and bipolar disorder. Science. 2018 Dec 14;362(6420):eaat8127. https://doi.org/10.1126/science.aat8127. PMID: 30545856; PMCID: PMC6443102. Gao L, Liu F, Hou PP, Manaenko A, Xiao ZP, Wang F, Xu TL, Hu Q. Neurons Release Injured Mitochondria as "Help-Me" Signaling After Ischemic Stroke. Front Aging Neurosci. 2022 Mar 3;14:785761. https://doi.org/10.3389/fnagi.2022.785761. PMID: 35309888; PMCID: PMC8926840. Gelot AB, Represa A. Progression of Fetal Brain Lesions in Tuberous Sclerosis Complex. Front Neurosci. 2020 Aug 21;14:899. https://doi.org/10.3389/fnins.2020.00899. PMID: 32973442; PMCID: PMC7472962. Gönczy P, Hatzopoulos GN. Centriole assembly at a glance. J Cell Sci. 2019 Feb 20;132(4):jcs228833. https://doi.org/10.1242/jcs.228833. PMID: 30787112. Greig LC, Woodworth MB, Galazo MJ, Padmanabhan H, Macklis JD. Molecular logic of neocortical projection neuron specification, development and diversity. Nat Rev Neurosci. 2013 Nov;14(11):755-69. https://doi.org/10.1038/nrn3586. Epub 2013 Oct 9. PMID: 24105342; PMCID: PMC3876965. Guerrini R, Dobyns WB. Malformations of cortical development: clinical features and genetic causes. Lancet Neurol. 2014 Jul;13(7):710-26. https://doi.org/10.1016/S1474-4422(14)70040-7. Epub 2014 Jun 2. PMID: 24932993; PMCID: PMC5548104. Guillén-Boixet J, Kopach A, Holehouse AS, Wittmann S, Jahnel M, et al. RNA-Induced Conformational Switching and Clustering of G3BP Drive Stress Granule Assembly by Condensation. Cell. 2020 Apr 16;181(2):346-361.e17. https://doi.org/10.1016/j.cell.2020.03.049. PMID: 32302572; PMCID: PMC7181197. Gumbs SBH, Berdenis van Berlekom A, Kübler R, Schipper PJ, Gharu L, Boks MP, Ormel PR, Wensing AMJ, de Witte LD, Nijhuis M. Characterization of HIV-1 Infection in Microglia-Containing Human Cerebral Organoids. Viruses. 2022 Apr 16;14(4):829. https://doi.org/10.3390/v14040829. PMID: 35458559; PMCID: PMC9032670. Halvorsen M, Gould L, Wang X, Grant G, Moya R, et al. De novo mutations in childhood cases of sudden unexplained death that disrupt intracellular Ca2+ regulation. Proc Natl Acad Sci U S A. 2021 Dec 28;118(52):e2115140118. https://doi.org/10.1073/pnas.2115140118. PMID: 34930847; PMCID: PMC8719874. Hansen DV, Lui JH, Flandin P, Yoshikawa K, Rubenstein JL, Alvarez-Buylla A, Kriegstein AR. Non-epithelial stem cells and cortical interneuron production in the human ganglionic eminences. Nat Neurosci. 2013 Nov;16(11):1576-87. https://doi.org/10.1038/nn.3541. Epub 2013 Oct 6. PMID: 24097039; PMCID: PMC4191718. Harowitz J, Crandall L, McGuone D, Devinsky O. Seizure-related deaths in children: The expanding spectrum. Epilepsia. 2021 Mar;62(3):570-582. https://doi.org/10.1111/epi.16833. Epub 2021 Feb 14. PMID: 33586153; PMCID: PMC7986159. Hayakawa K, Esposito E, Wang X, Terasaki Y, Liu Y, Xing C, Ji X, Lo EH. Transfer of mitochondria from astrocytes to neurons after stroke. Nature. 2016 Jul 28;535(7613):551-5. https://doi.org/10.1038/nature18928. Erratum in: Nature. 2016 Sep 14;539(7627):123. PMID: 27466127; PMCID: PMC4968589. Hefti MM, Kinney HC, Cryan JB, Haas EA, Chadwick AE, Crandall LA, Trachtenberg FL, Armstrong DD, Grafe M, Krous HF. Sudden unexpected death in early childhood: general observations in a series of 151 cases: Part 1 of the investigations of the San Diego SUDC Research Project. Forensic Sci Med Pathol. 2016 Mar;12(1):4-13. https://doi.org/10.1007/s12024-015-9724-2. Epub 2016 Jan 19. PMID: 26782961; PMCID: PMC4752958. Henske EP, Jóźwiak S, Kingswood JC, Sampson JR, Thiele EA. Tuberous sclerosis complex. Nat Rev Dis Primers. 2016 May 26;2:16035. https://doi.org/10.1038/nrdp.2016.35. PMID: 27226234. Henske EP, Scheithauer BW, Short MP, Wollmann R, Nahmias J, Hornigold N, van Slegtenhorst M, Welsh CT, Kwiatkowski DJ. Allelic loss is frequent in tuberous sclerosis kidney lesions but rare in brain lesions. Am J Hum Genet. 1996 Aug;59(2):400-6. PMID: 8755927; PMCID: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1914733. Hesdorffer DC, Crandall LA, Friedman D, Devinsky O. Sudden unexplained death in childhood: A comparison of cases with and without a febrile seizure history. Epilepsia. 2015 Aug;56(8):1294-300. https://doi.org/10.1111/epi.13066. Epub 2015 Jun 29. PMID: 26120007. Hessami K, Norooznezhad AH, Monteiro S, Barrozo ER, Abdolmaleki AS, Arian SE, Zargarzadeh N, Shekerdemian LS, Aagaard KM, Shamshirsaz AA. COVID-19 Pandemic and Infant Neurodevelopmental Impairment: A Systematic Review and Meta-analysis. JAMA Netw Open. 2022 Oct 3;5(10):e2238941. https://doi.org/10.1001/jamanetworkopen.2022.38941. PMID: 36306133; PMCID: PMC9617178. Hodge RD, Bakken TE, Miller JA, Smith KA, Barkan ER, et al. Conserved cell types with divergent features in human versus mouse cortex. Nature. 2019 Sep;573(7772):61-68. https://doi.org/10.1038/s41586-019-1506-7. Epub 2019 Aug 21. PMID: 31435019; PMCID: PMC6919571. Holm IA, Poduri A, Crandall L, Haas E, Grafe MR, Kinney HC, Krous HF. Inheritance of febrile seizures in sudden unexplained death in toddlers. Pediatr Neurol. 2012 Apr;46(4):235-9. https://doi.org/10.1016/j.pediatrneurol.2012.02.007. PMID: 22490769; PMCID: PMC4009678. Jabari S, Kobow K, Pieper T, Hartlieb T, Kudernatsch M, et al. DNA methylation-based classification of malformations of cortical development in the human brain. Acta Neuropathol. 2022 Jan;143(1):93-104. https://doi.org/10.1007/s00401-021-02386-0. Epub 2021 Nov 19. PMID: 34797422; PMCID: PMC8732912. Jain S, Wheeler JR, Walters RW, Agrawal A, Barsic A, Parker R. ATPase-Modulated Stress Granules Contain a Diverse Proteome and Substructure. Cell. 2016 Jan 28;164(3):487-98. https://doi.org/10.1016/j.cell.2015.12.038. Epub 2016 Jan 14. PMID: 26777405; PMCID: PMC4733397. Jansen LA, Mirzaa GM, Ishak GE, O'Roak BJ, Hiatt JB, et al. PI3K/AKT pathway mutations cause a spectrum of brain malformations from megalencephaly to focal cortical dysplasia. Brain. 2015 Jun;138(Pt 6):1613-28. https://doi.org/10.1093/brain/awv045. Epub 2015 Feb 25. PMID: 25722288; PMCID: PMC4614119. Jia X, Zhang S, Tan S, Du B, He M, et al. De novo variants in genes regulating stress granule assembly associate with neurodevelopmental disorders. Sci Adv. 2022 Aug 19;8(33):eabo7112. https://doi.org/10.1126/sciadv.abo7112. Epub 2022 Aug 17. PMID: 35977029; PMCID: PMC9385150. Kichula EA, Huntley GW. Developmental and comparative aspects of posterior medial thalamocortical innervation of the barrel cortex in mice and rats. J Comp Neurol. 2008 Jul 20;509(3):239-58. https://doi.org/10.1002/cne.21690. PMID: 18496871; PMCID: PMC4913357. Kiese K, Jablonski J, Hackenbracht J, Wrosch JK, Groemer TW, Kornhuber J, Blümcke I, Kobow K. Epigenetic control of epilepsy target genes contributes to a cellular memory of epileptogenesis in cultured rat hippocampal neurons. Acta Neuropathol Commun. 2017 Oct 31;5(1):79. https://doi.org/10.1186/s40478-017-0485-x. PMID: 29089052; PMCID: PMC5664434. Kinney DK, Miller AM, Crowley DJ, Huang E, Gerber E. Autism prevalence following prenatal exposure to hurricanes and tropical storms in Louisiana. J Autism Dev Disord. 2008 Mar;38(3):481-8. https://doi.org/10.1007/s10803-007-0414-0. Epub 2007 Jul 6. PMID: 17619130. Kinney HC, Poduri AH, Cryan JB, Haynes RL, Teot L, et al. Hippocampal Formation Maldevelopment and Sudden Unexpected Death across the Pediatric Age Spectrum. J Neuropathol Exp Neurol. 2016 Oct;75(10):981-997. https://doi.org/10.1093/jnen/nlw075. Epub 2016 Sep 9. PMID: 27612489; PMCID: PMC6281079. Kishi M, Pan YA, Crump JG, Sanes JR. Mammalian SAD kinases are required for neuronal polarization. Science. 2005 Feb 11;307(5711):929-32. https://doi.org/10.1126/science.1107403. PMID: 15705853. Kitahara T, Sakaguchi H, Morizane A, Kikuchi T, Miyamoto S, Takahashi J. Axonal Extensions along Corticospinal Tracts from Transplanted Human Cerebral Organoids. Stem Cell Reports. 2020 Aug 11;15(2):467-481. https://doi.org/10.1016/j.stemcr.2020.06.016. Epub 2020 Jul 16. PMID: 32679062; PMCID: PMC7419717. Kobow K, Blümcke I. The emerging role of DNA methylation in epileptogenesis. Epilepsia. 2012 Dec;53 Suppl 9:11-20. https://doi.org/10.1111/epi.12031. PMID: 23216575. Kobow K, Jabari S, Pieper T, Kudernatsch M, Polster T, et al. Mosaic trisomy of chromosome 1q in human brain tissue associates with unilateral polymicrogyria, very early-onset focal epilepsy, and severe developmental delay. Acta Neuropathol. 2020 Dec;140(6):881-891. https://doi.org/10.1007/s00401-020-02228-5. Epub 2020 Sep 26. PMID: 32979071; PMCID: PMC7666281. Kobow K, Kaspi A, Harikrishnan KN, Kiese K, Ziemann M, et al. Deep sequencing reveals increased DNA methylation in chronic rat epilepsy. Acta Neuropathol. 2013 Nov;126(5):741-56. https://doi.org/10.1007/s00401-013-1168-8. Epub 2013 Sep 5. PMID: 24005891; PMCID: PMC3825532. Kobow K, Ziemann M, Kaipananickal H, Khurana I, Mühlebner A, et al. Genomic DNA methylation distinguishes subtypes of human focal cortical dysplasia. Epilepsia. 2019 Jun;60(6):1091-1103. https://doi.org/10.1111/epi.14934. Epub 2019 May 10. PMID: 31074842; PMCID: PMC6635741. Kon FC, Vázquez RZ, Lang A, Cohen MC. Hippocampal abnormalities and seizures: a 16-year single center review of sudden unexpected death in childhood, sudden unexpected death in epilepsy and SIDS. Forensic Sci Med Pathol. 2020 Sep;16(3):423-434. https://doi.org/10.1007/s12024-020-00268-7. Epub 2020 Jul 25. PMID: 32712908. Kuzniecky RI. MRI in cerebral developmental malformations and epilepsy. Magn Reson Imaging. 1995;13(8):1137-45. https://doi.org/10.1016/0730-725x(95)02024-n. PMID: 8750328. Kwan KY, Sestan N, Anton ES. Transcriptional co-regulation of neuronal migration and laminar identity in the neocortex. Development. 2012 May;139(9):1535-46. https://doi.org/10.1242/dev.069963. PMID: 22492350; PMCID: PMC3317962. Ledford H. Severe COVID could cause markers of old age in the brain. Nature. 2022 Dec;612(7940):389. https://doi.org/10.1038/d41586-022-04253-8. PMID: 36471141. Lee WS, Baldassari S, Stephenson SEM, Lockhart PJ, Baulac S, Leventer RJ. Cortical Dysplasia and the mTOR Pathway: How the Study of Human Brain Tissue Has Led to Insights into Epileptogenesis. Int J Mol Sci. 2022 Jan 25;23(3):1344. https://doi.org/10.3390/ijms23031344. PMID: 35163267; PMCID: PMC8835853. Leitner DF, McGuone D, William C, Faustin A, Askenazi M, et al. Blinded review of hippocampal neuropathology in sudden unexplained death in childhood reveals inconsistent observations and similarities to explained paediatric deaths. Neuropathol Appl Neurobiol. 2022 Feb;48(1):e12746. https://doi.org/10.1111/nan.12746. Epub 2021 Jul 16. PMID: 34164845; PMCID: PMC8777468. a Leitner DF, William C, Faustin A, Askenazi M, Kanshin E, et al. Proteomic differences in hippocampus and cortex of sudden unexplained death in childhood. Acta Neuropathol. 2022 May;143(5):585-599. https://doi.org/10.1007/s00401-022-02414-7. Epub 2022 Mar 25. PMID: 35333953; PMCID: PMC8953962.b Leventer RJ, Phelan EM, Coleman LT, Kean MJ, Jackson GD, Harvey AS. Clinical and imaging features of cortical malformations in childhood. Neurology. 1999 Sep 11;53(4):715-22. https://doi.org/10.1212/wnl.53.4.715. PMID: 10489031. Linaro D, Vermaercke B, Iwata R, Ramaswamy A, Libé-Philippot B, et al. Xenotransplanted Human Cortical Neurons Reveal Species-Specific Development and Functional Integration into Mouse Visual Circuits. Neuron. 2019 Dec 4;104(5):972-986.e6. https://doi.org/10.1016/j.neuron.2019.10.002. Epub 2019 Nov 21. PMID: 31761708; PMCID: PMC6899440. Liu B, Qian SB. Translational reprogramming in cellular stress response. Wiley Interdiscip Rev RNA. 2014 May-Jun;5(3):301-15. https://doi.org/10.1002/wrna.1212. Epub 2013 Dec 23. PMID: 24375939; PMCID: PMC3991730. Mansour AA, Gonçalves JT, Bloyd CW, Li H, Fernandes S, Quang D, Johnston S, Parylak SL, Jin X, Gage FH. An in vivo model of functional and vascularized human brain organoids. Nat Biotechnol. 2018 Jun;36(5):432-441. https://doi.org/10.1038/nbt.4127. Epub 2018 Apr 16. PMID: 29658944; PMCID: PMC6331203. Markmiller S, Soltanieh S, Server KL, Mak R, Jin W, et al. Context-Dependent and Disease-Specific Diversity in Protein Interactions within Stress Granules. Cell. 2018 Jan 25;172(3):590-604.e13. https://doi.org/10.1016/j.cell.2017.12.032. PMID: 29373831; PMCID: PMC5969999. Martin KC, Ephrussi A. mRNA localization: gene expression in the spatial dimension. Cell. 2009 Feb 20;136(4):719-30. https://doi.org/10.1016/j.cell.2009.01.044. PMID: 19239891; PMCID: PMC2819924. McGuone D, Leitner D, William C, Faustin A, Leelatian N, et al. Neuropathologic Changes in Sudden Unexplained Death in Childhood. J Neuropathol Exp Neurol. 2020 Mar 1;79(3):336-346. https://doi.org/10.1093/jnen/nlz136. PMID: 31995186; PMCID: PMC7036658. Meisler MH, Kearney JA. Sodium channel mutations in epilepsy and other neurological disorders. J Clin Invest. 2005 Aug;115(8):2010-7. https://doi.org/10.1172/JCI25466. PMID: 16075041; PMCID: PMC1180547. Miyata T, Kawaguchi D, Kawaguchi A, Gotoh Y. Mechanisms that regulate the number of neurons during mouse neocortical development. Curr Opin Neurobiol. 2010 Feb;20(1):22-8. https://doi.org/10.1016/j.conb.2010.01.001. PMID: 20138502. Mizuguchi M. Abnormal giant cells in the cerebral lesions of tuberous sclerosis complex. Congenit Anom (Kyoto). 2007 Mar;47(1):2-8. https://doi.org/10.1111/j.1741-4520.2006.00134.x. PMID: 17300684. Murphy JV, Craig L. Leigh's disease: significance of the biochemical changes in brain. J Neurol Neurosurg Psychiatry. 1975 Nov;38(11):1100-3. https://doi.org/10.1136/jnnp.38.11.1100. PMID: 1206418; PMCID: PMC492163. Nakanishi K, Niida H, Tabata H, Ito T, Hori Y, et al. Isozyme-Specific Role of SAD-A in Neuronal Migration During Development of Cerebral Cortex. Cereb Cortex. 2019 Aug 14;29(9):3738-3751. https://doi.org/10.1093/cercor/bhy253. PMID: 30307479; PMCID: PMC7335017. Narula N, Tester DJ, Paulmichl A, Maleszewski JJ, Ackerman MJ. Post-mortem Whole exome sequencing with gene-specific analysis for autopsy-negative sudden unexplained death in the young: a case series. Pediatr Cardiol. 2015 Apr;36(4):768-78. https://doi.org/10.1007/s00246-014-1082-4. Epub 2014 Dec 13. PMID: 25500949; PMCID: PMC4907366. O'Neill AC, Uzbas F, Antognolli G, Merino F, Draganova K, et al. Spatial centrosome proteome of human neural cells uncovers disease-relevant heterogeneity. Science. 2022 Jun 17;376(6599):eabf9088. https://doi.org/10.1126/science.abf9088. Epub 2022 Jun 17. PMID: 35709258. Ormel PR, Vieira de Sá R, van Bodegraven EJ, Karst H, Harschnitz O, et al. Microglia innately develop within cerebral organoids. Nat Commun. 2018 Oct 9;9(1):4167. https://doi.org/10.1038/s41467-018-06684-2. PMID: 30301888; PMCID: PMC6177485. Panagiotakos G, Pasca SP. A matter of space and time: Emerging roles of disease-associated proteins in neural development. Neuron. 2022 Jan 19;110(2):195-208. https://doi.org/10.1016/j.neuron.2021.10.035. Epub 2021 Nov 29. PMID: 34847355; PMCID: PMC8776599. Paredes MF, James D, Gil-Perotin S, Kim H, Cotter JA, et al. Extensive migration of young neurons into the infant human frontal lobe. Science. 2016 Oct 7;354(6308):aaf7073. https://doi.org/10.1126/science.aaf7073. PMID: 27846470; PMCID: PMC5436574. Parikshak NN, Swarup V, Belgard TG, Irimia M, Ramaswami G, et al. Genome-wide changes in lncRNA, splicing, and regional gene expression patterns in autism. Nature. 2016 Dec 15;540(7633):423-427. https://doi.org/10.1038/nature20612. Epub 2016 Dec 5. Erratum in: Nature. 2018 Aug;560(7718):E30. PMID: 27919067; PMCID: PMC7102905. Piel M, Meyer P, Khodjakov A, Rieder CL, Bornens M. The respective contributions of the mother and daughter centrioles to centrosome activity and behavior in vertebrate cells. J Cell Biol. 2000 Apr 17;149(2):317-30. https://doi.org/10.1083/jcb.149.2.317. PMID: 10769025; PMCID: PMC2175166. Qin W, Chan JA, Vinters HV, Mathern GW, Franz DN, Taillon BE, Bouffard P, Kwiatkowski DJ. Analysis of TSC cortical tubers by deep sequencing of TSC1, TSC2 and KRAS demonstrates that small second-hit mutations in these genes are rare events. Brain Pathol. 2010 Nov;20(6):1096-105. https://doi.org/10.1111/j.1750-3639.2010.00416.x. Epub 2010 Jul 13. PMID: 20633017; PMCID: PMC2951479. Raj B, Blencowe BJ. Alternative Splicing in the Mammalian Nervous System: Recent Insights into Mechanisms and Functional Roles. Neuron. 2015 Jul 1;87(1):14-27. https://doi.org/10.1016/j.neuron.2015.05.004. PMID: 26139367. Raju CS, Spatazza J, Stanco A, Larimer P, Sorrells SF, et al. Secretagogin is Expressed by Developing Neocortical GABAergic Neurons in Humans but not Mice and Increases Neurite Arbor Size and Complexity. Cereb Cortex. 2018 Jun 1;28(6):1946-1958. https://doi.org/10.1093/cercor/bhx101. PMID: 28449024; PMCID: PMC6019052. Ramaswami G, Won H, Gandal MJ, Haney J, Wang JC, Wong CCY, Sun W, Prabhakar S, Mill J, Geschwind DH. Integrative genomics identifies a convergent molecular subtype that links epigenomic with transcriptomic differences in autism. Nat Commun. 2020 Sep 25;11(1):4873. https://doi.org/10.1038/s41467-020-18526-1. PMID: 32978376; PMCID: PMC7519165. Real R, Peter M, Trabalza A, Khan S, Smith MA, et al. In vivo modeling of human neuron dynamics and Down syndrome. Science. 2018 Nov 16;362(6416):eaau1810. https://doi.org/10.1126/science.aau1810. Epub 2018 Oct 11. PMID: 30309905; PMCID: PMC6570619. Revah O, Gore F, Kelley KW, Andersen J, Sakai N, et al. Maturation and circuit integration of transplanted human cortical organoids. Nature. 2022 Oct;610(7931):319-326. https://doi.org/10.1038/s41586-022-05277-w. Epub 2022 Oct 12. PMID: 36224417; PMCID: PMC9556304. Roy A, Millen KJ, Kapur RP. Hippocampal granule cell dispersion: a non-specific finding in pediatric patients with no history of seizures. Acta Neuropathol Commun. 2020 Apr 21;8(1):54. https://doi.org/10.1186/s40478-020-00928-3. PMID: 32317027; PMCID: PMC7171777. Rubenstein JL, Rakic P. Genetic control of cortical development. Cereb Cortex. 1999 Sep;9(6):521-3. https://doi.org/10.1093/cercor/9.6.521. PMID: 10498269. Ruppe V, Dilsiz P, Reiss CS, Carlson C, Devinsky O, Zagzag D, Weiner HL, Talos DM. Developmental brain abnormalities in tuberous sclerosis complex: a comparative tissue analysis of cortical tubers and perituberal cortex. Epilepsia. 2014 Apr;55(4):539-50. https://doi.org/10.1111/epi.12545. Epub 2014 Feb 11. PMID: 24512506. Sahm F, Schrimpf D, Stichel D, Jones DTW, Hielscher T, et al. DNA methylation-based classification and grading system for meningioma: a multicentre, retrospective analysis. Lancet Oncol. 2017 May;18(5):682-694. https://doi.org/10.1016/S1470-2045(17)30155-9. Epub 2017 Mar 15. PMID: 28314689. Schurr J, Coras R, Rössler K, Pieper T, Kudernatsch M, et al. Mild Malformation of Cortical Development with Oligodendroglial Hyperplasia in Frontal Lobe Epilepsy: A New Clinico-Pathological Entity. Brain Pathol. 2017 Jan;27(1):26-35. https://doi.org/10.1111/bpa.12347. Epub 2016 Feb 22. PMID: 26748554; PMCID: PMC8029051. Smith RS, Kenny CJ, Ganesh V, Jang A, Borges-Monroy R, et al. Sodium Channel SCN3A (NaV1.3) Regulation of Human Cerebral Cortical Folding and Oral Motor Development. Neuron. 2018 Sep 5;99(5):905-913.e7. https://doi.org/10.1016/j.neuron.2018.07.052. Epub 2018 Aug 23. PMID: 30146301; PMCID: PMC6226006. Son G, Han J. Roles of mitochondria in neuronal development. BMB Rep. 2018 Nov;51(11):549-556. https://doi.org/10.5483/BMBRep.2018.51.11.226. PMID: 30269744; PMCID: PMC6283025. Sun W, Poschmann J, Cruz-Herrera Del Rosario R, Parikshak NN, Hajan HS, et al. Histone Acetylome-wide Association Study of Autism Spectrum Disorder. Cell. 2016 Nov 17;167(5):1385-1397.e11. https://doi.org/10.1016/j.cell.2016.10.031. PMID: 27863250. Takahara T, Maeda T. Transient sequestration of TORC1 into stress granules during heat stress. Mol Cell. 2012 Jul 27;47(2):242-52. https://doi.org/10.1016/j.molcel.2012.05.019. Epub 2012 Jun 21. PMID: 22727621. Taylor DC, Falconer MA, Bruton CJ, Corsellis JA. Focal dysplasia of the cerebral cortex in epilepsy. J Neurol Neurosurg Psychiatry. 1971 Aug;34(4):369-87. https://doi.org/10.1136/jnnp.34.4.369. PMID: 5096551; PMCID: PMC493805. Thiele EA. Managing and understanding epilepsy in tuberous sclerosis complex. Epilepsia. 2010 Feb;51 Suppl 1:90-1. https://doi.org/10.1111/j.1528-1167.2009.02458.x. PMID: 20331728. Vineethakumari C, Lüders J. Microtubule Anchoring: Attaching Dynamic Polymers to Cellular Structures. Front Cell Dev Biol. 2022 Mar 3;10:867870. https://doi.org/10.3389/fcell.2022.867870. PMID: 35309944; PMCID: PMC8927778. Voineagu I, Wang X, Johnston P, Lowe JK, Tian Y, Horvath S, Mill J, Cantor RM, Blencowe BJ, Geschwind DH. Transcriptomic analysis of autistic brain reveals convergent molecular pathology. Nature. 2011 May 25;474(7351):380-4. https://doi.org/10.1038/nature10110. PMID: 21614001; PMCID: PMC3607626. Walker RL, Ramaswami G, Hartl C, Mancuso N, Gandal MJ, de la Torre-Ubieta L, Pasaniuc B, Stein JL, Geschwind DH. Genetic Control of Expression and Splicing in Developing Human Brain Informs Disease Mechanisms. Cell. 2019 Oct 17;179(3):750-771.e22. https://doi.org/10.1016/j.cell.2019.09.021. Erratum in: Cell. 2020 Apr 16;181(2):484. Erratum in: Cell. 2020 Apr 30;181(3):745. PMID: 31626773; PMCID: PMC8963725. Wamsley B, Jaglin XH, Favuzzi E, Quattrocolo G, Nigro MJ, Yusuf N, Khodadadi-Jamayran A, Rudy B, Fishell G. Rbfox1 Mediates Cell-type-Specific Splicing in Cortical Interneurons. Neuron. 2018 Nov 21;100(4):846-859.e7. https://doi.org/10.1016/j.neuron.2018.09.026. Epub 2018 Oct 11. PMID: 30318414; PMCID: PMC6541232. Wang R, Zhang H, Du J, Xu J. Heat resilience in embryonic zebrafish revealed using an in vivo stress granule reporter. J Cell Sci. 2019 Oct 18;132(20):jcs234807. https://doi.org/10.1242/jcs.234807. PMID: 31558681; PMCID: PMC6826007. Wedderburn CJ, Weldon E, Bertran-Cobo C, Rehman AM, Stein DJ, Gibb DM, Yeung S, Prendergast AJ, Donald KA. Early neurodevelopment of HIV-exposed uninfected children in the era of antiretroviral therapy: a systematic review and meta-analysis. Lancet Child Adolesc Health. 2022 Jun;6(6):393-408. https://doi.org/10.1016/S2352-4642(22)00071-2. Epub 2022 Apr 26. PMID: 35483380; PMCID: PMC9090907. Wefers AK, Stichel D, Schrimpf D, Coras R, Pages M, et al. Isomorphic diffuse glioma is a morphologically and molecularly distinct tumour entity with recurrent gene fusions of MYBL1 or MYB and a benign disease course. Acta Neuropathol. 2020 Jan;139(1):193-209. https://doi.org/10.1007/s00401-019-02078-w. Epub 2019 Sep 28. PMID: 31563982; PMCID: PMC7477753. Wilsch-Bräuninger M, Huttner WB. Primary Cilia and Centrosomes in Neocortex Development. Front Neurosci. 2021 Oct 21;15:755867. https://doi.org/10.3389/fnins.2021.755867. PMID: 34744618; PMCID: PMC8566538. Wu YE, Parikshak NN, Belgard TG, Geschwind DH. Genome-wide, integrative analysis implicates microRNA dysregulation in autism spectrum disorder. Nat Neurosci. 2016 Nov;19(11):1463-1476. https://doi.org/10.1038/nn.4373. Epub 2016 Aug 29. PMID: 27571009; PMCID: PMC5841760. Xiong M, Tao Y, Gao Q, Feng B, Yan W, et al. Human Stem Cell-Derived Neurons Repair Circuits and Restore Neural Function. Cell Stem Cell. 2021 Jan 7;28(1):112-126.e6. https://doi.org/10.1016/j.stem.2020.08.014. Epub 2020 Sep 22. PMID: 32966778; PMCID: PMC7796915. Yang P, Mathieu C, Kolaitis RM, Zhang P, Messing J, et al. G3BP1 Is a Tunable Switch that Triggers Phase Separation to Assemble Stress Granules. Cell. 2020 Apr 16;181(2):325-345.e28. https://doi.org/10.1016/j.cell.2020.03.046. PMID: 32302571; PMCID: PMC7448383. Youn JY, Dunham WH, Hong SJ, Knight JDR, Bashkurov M, et al. High-Density Proximity Mapping Reveals the Subcellular Organization of mRNA-Associated Granules and Bodies. Mol Cell. 2018 Feb 1;69(3):517-532.e11. https://doi.org/10.1016/j.molcel.2017.12.020. Epub 2018 Jan 25. PMID: 29395067. Yu FH, Mantegazza M, Westenbroek RE, Robbins CA, Kalume F, Burton KA, Spain WJ, McKnight GS, Scheuer T, Catterall WA. Reduced sodium current in GABAergic interneurons in a mouse model of severe myoclonic epilepsy in infancy. Nat Neurosci. 2006 Sep;9(9):1142-9. https://doi.org/10.1038/nn1754. Epub 2006 Aug 20. Erratum in: Nat Neurosci. 2007 Jan;10(1):134. PMID: 16921370.
Copyright: © 2023 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |