|
|
|
Free Neuropathology 3:15 (2022) |
|
Review |
|
Neurovascular disease: 2022 update |
|
Louise D. McCullough |
|
Department of Neurology, McGovern Medical School, UTHealth Houston and Memorial Hermann Hospital, Houston, Texas, USA |
|
Corresponding author: |
|
Submitted: 25 April 2022 Accepted: 07 June 2022 Copyedited by: Félicia Jeannelle Published: 14 June 2022 |
|
Keywords: Stroke, Vascular malformations, Microglia, T cells, Neuroinflammation |
|
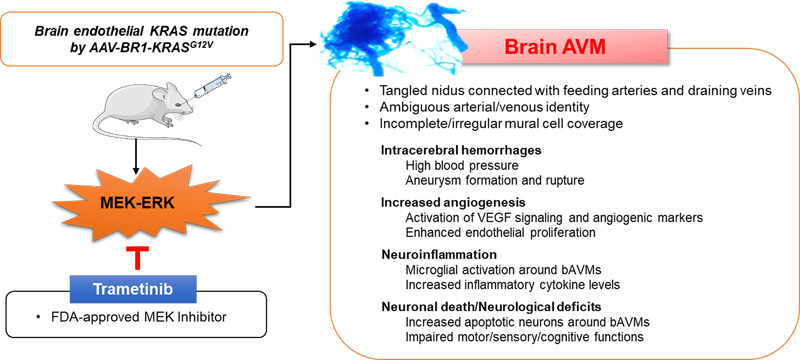
Abstract In this update we present a series of papers focused on topics that have emerged in vascular disease over the prior year. The first two papers focus on the pathogenesis of vascular malformations, the first on brain arteriovenous malformations, and the second on cerebral cavernous malformations. These disorders can lead to significant brain injuries from intracerebral hemorrhage (if they rupture) or other neurological complications, including seizures. The next set of papers reflects work that has advanced our understanding of how the brain and the immune system “communicate” after brain injury, including stroke (papers 3-6). The first of these shows that T cells are involved in white matter repair after ischemic injury, an effect dependent on microglia, demonstrating the important cross-talk between innate and adaptive immunity. The next two papers focus on B cells, which have been relatively understudied in the context of brain injury. The contribution of antigen-experienced B cells from the meninges and skull bone marrow, rather than blood-derived B cells in neuroinflammation opens up a very novel area of investigation. The possibility that antibody secreting B cells may contribute to vascular dementia will certainly be an active area for future investigations. Similarly, in paper 6, investigators found that CNS-infiltrating myeloid cells can originate from brain borders tissues. These cells have unique transcriptional signatures that are distinct from their blood-derived counterparts, and likely contribute to myeloid cell infiltration from bone-marrow niches in close proximity to the brain. The contribution of microglia, the primary innate immune cell of the brain, to amyloid deposition and propagation is then discussed, followed by work on how perivascular Aβ is potentially cleared along the cerebral vessels in patients with cerebral amyloid angiopathy. The final two papers focus on the contribution of senescent endothelial cells and pericytes. The first used a model of accelerated senescence (Hutchinson-Gilford progeria syndrome; HGPS) and shows the translational potential of an approach to reduce telomere shortening to slow aging. The final paper demonstrates how capillary pericytes contribute to basal blood flow resistance and slow modulation of blood flow throughout the brain. Interestingly, several of the papers identified therapeutic strategies that could be potentially translated into clinical populations. Introduction This “Neurovascular disease: 2022 update” presents topics that were selected as the top 10 papers and discoveries published in the field of neurovascular disorders in 2021. These papers were selected based on their broad appeal, those with new methods or discoveries, and those that have a direct link to neurovascular inflammation, vascular abnormalities and neuroimmunology. It is important to note that these top ten papers were subjectively selected by many individuals when queried regarding what papers they felt were most impactful in 2021 (individual contributors are listed in the acknowledgement section). Thus, there is certainly considerable “selection bias”. These papers were chosen by a large group of neurosurgery and neurology faculty, and are not ranked in any specific order or by number of citations. Overall, the theme of the selected papers reflects our growing understanding of how immunological events, both in the brain and in the periphery, alter the response to vascular and degenerative diseases. The field of neuroimmunology has seen tremendous growth over the past decade, and this is reflected in the papers selected. We apologize to all the authors we could not highlight due to space limitations. Topic 1: Selective endothelial hyperactivation of oncogenic KRAS induces brain arteriovenous malformations in mice Brain arteriovenous malformations (bAVMs) are a challenging cerebrovascular disease, characterized by tangled/enlarged vessels occurred by direct connections between arteries and veins without intervening capillary bed. bAVMs are a major cause of the most debilitating spontaneous intracerebral hemorrhage in children and young adults. However, the pathogenesis of the disease is largely unknown, and the treatment options for bAVM patients are severely limited mostly relying on risky surgical methods. Recently, a clinical study has reported that somatic activating KRAS mutations were detected in the endothelium of ~60% of human sporadic bAVMs (1). In addition, mosaic variants have also been found in the BRAF and MAP2K1/MEK in intracranial or extracranial AVMs, especially in high-flow vascular malformations (2). These observations suggest the critical role of KRAS signaling, especially RAF/MEK/ERK kinase cascade in AVM pathogenesis. Based on this evidence, the authors tested the role of KRAS mutation in bAVM development using an adeno-associated virus (AAV)-BR1 specifically targeting brain vascular endothelial cells. Strikingly, the systemic administration of AAV-BR1 encoding human KRASG12V gene (AAV-BR1-KRASG12V) induced vascular malformations in the mouse brains that recapitulates salient features of human bAVMs including the tangled nidus connected with feeding arteries and draining veins, multi-focal spontaneous intracerebral hemorrhages, ambiguous arterial/venous identity and irregular/incomplete mural cell coverage, and enhanced proliferating endothelial cells and angiogenic factors. The accumulation of activated microglia/macrophages, enhanced inflammatory response, and neuronal death were observed around bAVMs, and mice with bAVM showed impaired behavioral functions. The study found the ERK activation, but not pAkt and p38, in the bAVMs suggesting that MEK/ERK signaling is the key mediator for the mutant KRAS-induced bAVM development, and finally revealed that Trametinib (an FDA-approved MEK inhibitor for cancer) attenuates the growth of KRASG12V-induced bAVMs (Figure 1). The novel approach by Park et al. (3) clearly confirmed the causative role of KRAS mutations in bAVM development, and generated a novel/clinically-relevant mouse model of bAVMs that will be useful to define the underlying pathomechanism in bAVMs. This is the first preclinical study showing the efficacy of pharmacological intervention in sporadic bAVMs associated with KRAS mutation providing a potential therapeutic strategy for bAVM patients by repurposing a cancer drug.
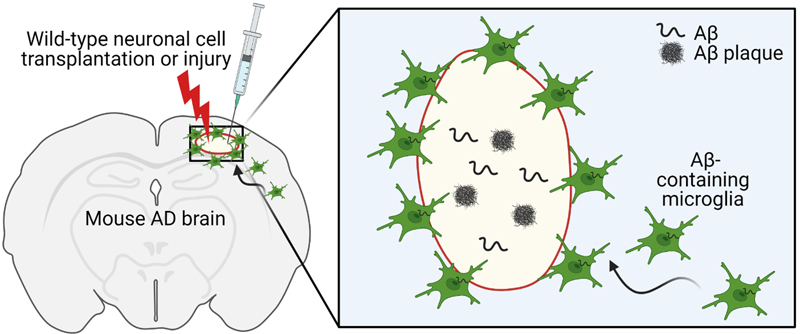
Figure 1. The characteristics of mouse bAVMs induced by brain endothelial KRASG12V overexpression. Systemic injection of AAV-BR1 carrying KRASG12V (AAV-BR1-KRASG12V)-induced abnormal vasculatures that recapitulate the salient features of human bAVMs including tangled nidus, irregular mural cell coverages, and spontaneous hemorrhages. Enhanced endogenous vascular endothelial growth factor (VEGF) signaling and endothelial cell proliferation were observed in the bAVMs. Accumulating activated microglia were seen around bAVMs and accompanied with increased expression of inflammatory factors (e.g., cytokines, matrix metalloproteinases) and apoptotic neurons. Mice with bAVM showed significantly impaired behavioral function. Prolonged treatment with trametinib, an FDA-approved MEK inhibitor (for six weeks from one day after the AAV-BR1-KRASG12V) inhibited bAVM growth suggesting the MEK/ERK signaling is critical in bAVM development induced by KRAS mutation (Park et al., 2021). Topic 2: PIK3CA and CCM mutations fuel cavernomas through a cancer-like mechanism Another interesting work, also on the topic of brain vascular malformations, comes from the laboratory of Dr. Khan (4). In this work the authors focused on cerebral cavernous malformations (CCMs) rather than the AVMs explored in the prior manuscript. CCMs develop due to inactivation of the endothelial CCM protein complex, which is needed to decrease the activity of the kinase MEKK3. These malformations can lead to intracerebral hemorrhages and seizures, and often affect younger individuals. The clinical spectrum of disease presentation, including growth and rupture, varies between individuals. Some CCMs are clinically silent, whereas others grow rapidly and are at risk for repeated hemorrhages. Identifying which subset of CCMs are more likely to become symptomatic is important, as surgical resection may be warranted. However, surgery has its own risks, and patients often have multiple lesions. Genetic studies have implicated a monogenic basis for CCM disease associated with biallelic loss of function mutations in three genes, KRIT1, CCM2, and PDCD10, that encode the components of a heterotrimeric CCM protein complex, as described by others. In this work, the authors demonstrate that the growth of CCMs requires increased signaling through the phosphatidylinositol-3-kinase (PI3K)-mTOR pathway as well as loss of function of the CCM complex. This work identified that symptomatic CCM disease arises through a cancer-like program, with the accumulation of multiple somatic mutations in the same cell. This results in both the loss of a vascular malformation suppressor gene (i.e., the CCM gene) and the gain of vascular malformation growth gene (i.e. PIK3CA). The authors found somatic gain-of-function mutations in PIK3CA and loss-of-function mutations in the CCM complex in the same cells in the majority of human CCMs. They then moved to mouse models and found that the growth of CCMs requires both PI3K gain of function (GOF) and CCM loss of function (LOF) in endothelial cells, as both were required for CCM development. Both CCM loss of function and increased expression of the transcription factor KLF4 (a downstream effector of MEKK3) increased mTOR signaling in endothelial cells. Importantly, the authors then went on to utilize Rapamycin, a mTORC1 inhibitor in mice. As single dose given at P2 reduced CCM formation by 75%. In adult mice, chronic administration of Rapamycin effectively blocked the formation of CCMs in mouse models. This work suggests that there is a “triple-hit” mechanism involving the acquisition of as many as three distinct genetic mutations that culminate in CCM LOF and PIK3CA GOF and contribute to rapidly growing, clinically symptomatic CCMs. This three-hit mechanism is analogous to cancer, in which aggressive vascular malformations arise through both the loss of vascular “suppressor genes” that limit vessel growth and gain of function of a vascular “oncogene” that then stimulates excess vascular growth. Importantly, these findings suggest that aggressive CCMs may be treatable using clinically approved mTORC1 inhibitors. Topic 3: Treg cell-derived osteopontin promotes microglia-mediated white matter repair after ischemic stroke In this paper (5) the role of regulatory T cells (Treg) in stroke recovery was examined. Shi et al. sorted infiltrating immune cells from the ischemic brain at 5 and 14 days after transient middle cerebral artery occlusion (MCAO). The numbers of T cells increased robustly from 5 to 14 days after stroke, suggesting a possible role of these cells in stroke pathophysiology and recovery. The authors subclustered and more deeply assessed the 14-day T cell population. Seven subclusters were identified, including CD4+ Treg cells, CD4+ memory T cells, natural killer T (NKT) cells, macrophage-like T cells, and populations of CD8+ T effector cells. The distinct Treg cell subcluster expressed transcripts of canonical Treg cell markers such as Cd4, Il2ra (CD25), Foxp3, and Ikzf2 (Helios), which is essential for T cell regulatory function. Temporal changes in the CD3+ CD4+ CD25+- Foxp3+ Treg cell population were assessed in the ischemic brain using flow cytometry at different time points (3, 5, 7, 14, and 35 days). Minimal Treg cell infiltration was observed 3 and 5 days after MCAO. The number of infiltrating Treg cells significantly increased from 7 days onward, increasing until at least 35 days post stroke. The visualization of t-distributed stochastic neighbor embedding (viSNE) map of flow cytometry data revealed a prominent Treg cell population among all CD4+ T cells in single-cell brain suspensions collected 14 days after stroke. These cells expressed CD25 and Foxp3. Consistent with recent studies, the majority of brain-infiltrating Treg cells were found to be Helios+, a population recently identified as phenotypically active Treg cells. These cells accumulated near white matter. Higher percentages of infiltrating Treg cells in the ischemic brain expressed osteopontin (OPN) compared with circulating Treg cells from sham or stroke mice. Bioinformatic analyses predicted strong interactions between Treg cells and microglia through OPN and integrin receptors, which were then confirmed experimentally by a series of in vivo and in vitro studies. The authors then selectively depleted Tregs using a diphtheria toxin method and found that loss of these cells impaired motor recovery and reduced myelin coverage, which was confirmed by whole brain ex vivo diffusion tensor imaging and electron microscopy. The beneficial effects of Treg cells were linked to oligodendrogenesis that was dependent on microglia. Microglia depletion, but not T cell lymphopenia, mitigated the beneficial effects of transferred Treg cells on white matter regeneration. Mechanistically, Treg cell-derived osteopontin acted through integrin receptors on microglia to enhance microglial reparative activity, consequently promoting oligodendrogenesis and white matter repair. These results confirm that Treg cell-microglia interactions enhance a reparative microglial phenotype that promotes OPC differentiation. Therefore, Treg cells appear to contribute to brain repair via both indirect immunomodulation of microglial responses as well as direct trophic effects. Boosting Treg cell numbers could be a practical approach to improve WM repair and functional recovery after stroke. Topic 4: Heterogeneity of meningeal B cells reveals a lymphopoietic niche at the CNS borders Meningeal immune cells are thought to originate from the systemic blood circulation. However, recent findings of developmentally immature B cells in the mouse central nervous system question this assumption. Brioschi et al. (6) used single cell analysis from parabiotic and chimeric mouse models to demonstrate that the skull bone marrow contains a hematopoietic niche from which extravascular meningeal B cells can arise to participate in the local immune response. Meningeal B cells are capable of trafficking through meningeal lymphatics to the cervical lymph nodes, enabling them to participate in antigen presentation linking brain and systemic immunity. These meningeal B cells are composed of a heterogeneous population with respect to their developmental and maturity stage, with late (B220lowCD43low), and mature (B220highCD43−) subsets representing over 80% of the total B cell population. This was confirmed by single cell RNA sequencing (scRNA-seq) and CyTOF mass spectrometry. scRNA-seq analysis showed that the meningeal and skull bone marrow B cells exhibited overlapping transcriptomic phenotypes, and that these were distinct from the blood-sourced B cells. Targeted gamma irradiation of the skull followed by reconstitution with donor bone marrow cells was used to generate a skull bone marrow transplantation model to investigate the origin of the meningeal B cells. The skull bone marrow chimera experiments confirmed that the meningeal B cells originate from the skull. Taken together, this study showed that skull bone marrow provides an accessible source of B cells that are locally educated by CNS-derived antigens. Lastly, scRNAseq data showed an age-dependent accumulation of blood-derived antigen-experienced B cells (identified as B220highCD23−CD2+Sca1+ cells) in the meninges. Meninges from aged animals (20-25 months of age) housed a significant population of IgM+ and IgG+ plasma cells, indicating their antigen-experienced status. These specific “age-associated B cells” (ABCs) accumulating in the meninges with aging were found to originate from the peripheral circulation, and represent a separate pool of B cells from those derived from the calvaria. Given the importance of B cells in the regulation of neuroinflammation and autoimmunity, meningeal B cells may be a critical subset of the adaptive immune cells, uniquely positioned to contribute to the pool of self-reactive B cells seen with aging (see next paper). Topic 5: Single-cell profiling of CNS border compartment leukocytes reveals that B cells and their progenitors reside in non-diseased meninges Central nervous system (CNS) meninges (dura, arachnoid, and pia) surround the CNS tissue at its critical borders. Recent studies examining the dural lymphatics have highlighted the importance of these CNS interfaces and their immune landscape. Schafflick et al. (7) used scRNA-seq of sorted leukocytes from rat meninges, blood, and cerebrospinal fluid (CSF) to identify unique cell compositions within the dura. Intravenous immune labeling using fluorophore-labeled CD45 (common leukocyte marker) antibody was used to exclude intravascular immune cells. Comparing scRNA-seq data from different tissues revealed that the dura contained a large cluster of B cells. These dural B cells were subclustered into Cxcr4high (signifying immature bone marrow B cells) versus Ccl2highCxcl1/2highCcl4highItgb2high (signifying chemotactically active B cells). Using myelin oligodendrocyte glycoprotein (MOG)35–55 peptide-induced experimental autoimmune encephalomyelitis (EAE) models of neuroinflammation, the authors showed that inflammation drives phenotypic changes of dural B cells by favoring class-switched plasma cells with higher expression of IgA than IgG. Dural B cells exhibited higher MHC class II and other markers, indicating an increased antigen-presenting capacity under inflammatory conditions. Location of meningeal B cells under homeostatic conditions was then investigated using immunofluorescence of whole mount dura preparations after intravenous injections with a fluorophore-labeled CD45 antibody. Results showed that dural B cells are located both within and outside of blood and lymphatic vessels, and interestingly, the extravascular B cells were particularly abundant along the skull sutures. Next, the authors confirmed the presence of meningeal B cells in human autopsy samples using CD19+CD20+CD79A+ stainings, and found that human dura also houses a relevant population of B cells. Using parabiotic mouse models with different CD45 haplotypes, the precursor dural B cells were shown to be tissue resident with capacity for local proliferation, and the authors claimed their skull labeling approach challenges the presumptive skull-to-dura immune cell flux. This study provided elegant data regarding the presence of a previously-neglected, strategically-located population of meningeal B cells with high potency for antigen-presentation, chemotaxis, and regulation of CNS-antigen-specific neuroinflammation. Topic 6: Skull and vertebral bone marrow are myeloid cell reservoirs for the meninges and CNS parenchyma A third paper (8) that integrates well with the theme of the prior two, also examined skull and vertebral bone marrow, but this focused on the myeloid compartment. Meninges as evidenced from this triad of papers, contain a large pool of immune cells. However, unlike the prior papers, this paper focused on innate immune cells rather than adaptive B cells. Myeloid cells are some of the earlies responders to brain injury. It is well known that these cells can migrate from the periphery to invade the CNS. This paper shows that these are not only “blood-derived” but can also come into the CNS, rapidly, from the skull bone marrow. The perivascular spaces and the meningeal membranes that cover the CNS border zones are also populated by a variety of myeloid cells. In this work, Cugarra and others show that CNS-associated bone marrow niches in the skull and vertebrae are also myeloid reservoirs for the meninges and CNS parenchyma. Under homeostasis, these bone marrow pools supply the brain and spinal dural meninges with monocytes and neutrophils via direct dural-bone marrow connections. Using elegant fate mapping, dural Ly6C+ monocytes and neutrophils were suspected to be from a local source rather than the blood. A calvaria bone-flap with accompanying bone marrow reservoir from GFP+ mice was transplanted into wild-type (WT) mice with an intact dura. GFP+ cells including CCR2+ monocytes, IBA1+ macrophages, and CD31+ vasculature was seen in the underlying dura of the transplanted calvaria flap. Thus, under homeostatic conditions, the myeloid niche distributed along the brain borders receives a substantial input from the skull bone marrow, which appears to act as a critical dispenser of myeloid cells. This provides an example of a healthy tissue hosting myeloid cells that are continuously replenished by a source that does not use blood as a major route. The authors then went one step further to determine if these cells were involved in an early response to injury using parabiotic models in three injury paradigms, experimental autoimmune encephalitis, spinal cord injury, and optic nerve crush. After injury and in neuroinflammatory diseases, the authors found that these cells can mobilize to infiltrate the CNS parenchyma, and display distinct phenotypes from their blood-derived counterparts. CNS-infiltrating Ly6C+ monocytes (but not neutrophils, CD4 T cells, or Ly6C− monocytes) were supplied not from the blood, but by adjacent skull and vertebral bone marrow. It is becoming increasingly clear that under pathological conditions, CNS-infiltrating myeloid cells can originate from brain borders and display transcriptional signatures distinct from their blood-derived counterparts. These myeloid cells from bone-marrow niches in close proximity to the brain can supply innate immune cells under both homeostatic and pathological conditions. Topic 7: Microglia contribute to the propagation of Aβ into unaffected brain tissue This very recent study revealed that microglia may act as Aβ carriers in the brain of Alzheimer’s disease (AD) model mice (9). D’Errico and colleagues transplanted embryonic neuronal cells from WT mice into the neocortex of pre-symptomatic 5xFAD transgenic mice, a mouse model recapitulating AD-related pathology. Interestingly, they found that Aβ plaques developed as early as 4 weeks within the grafted neuronal cells and continually increase over time. They confirmed that this was not due to anterograde transport, as very few host processes entered the graft using Thy1-GFP/5xFAD mice, confirming that the spread of Aβ is independent of axonal transport. The greatest amyloid accumulation was along the graft border, an area with higher microglia density. Therefore, the authors next investigated whether host microglia are able to invade the WT graft using Cx3cr1+/−/5xFAD mice. Even as early as 2 weeks after transplantation, migration of host CX3CR1+ microglia from CX3CR1+/-/5xFAD mice was evident within the WT graft, indicating the ability of microglia to act as an Aβ carrier leading to graft invasion. Using a variety of methods, the authors confirmed that the microglia were not host derived, or peripheral infiltrating monocytes. They also confirmed using both in vitro and in vivo models that a decline in microglial phagocytosis of Aβ occurs in both old WT and 5xFAD mice. Several additional models including aged 5xFAD mice (with less microglial phagocytic activity), IRF8-/-/5xFAD mice (with less microglial branching), and microglia-depleted 5xFAD mice using CSF-1R inhibition exhibited a reduction of Aβ plaques in the WT grafts, further corroborating the specific role of microglia in regulating Aβ propagation. This work confirms that microglia are involved in the pathogenic spread of Aβ. They also went one step further in a final experiment. The authors used a model of laser-induced focal tissue injury and two-photon microscopy, and confirmed that Aβ can be transported by microglia and form plaques in the lesioned tissue (Figure 2). In conclusion, this study suggests a novel role of microglia in the propagation of Aβ pathology in neurodegenerative and neural injury models.
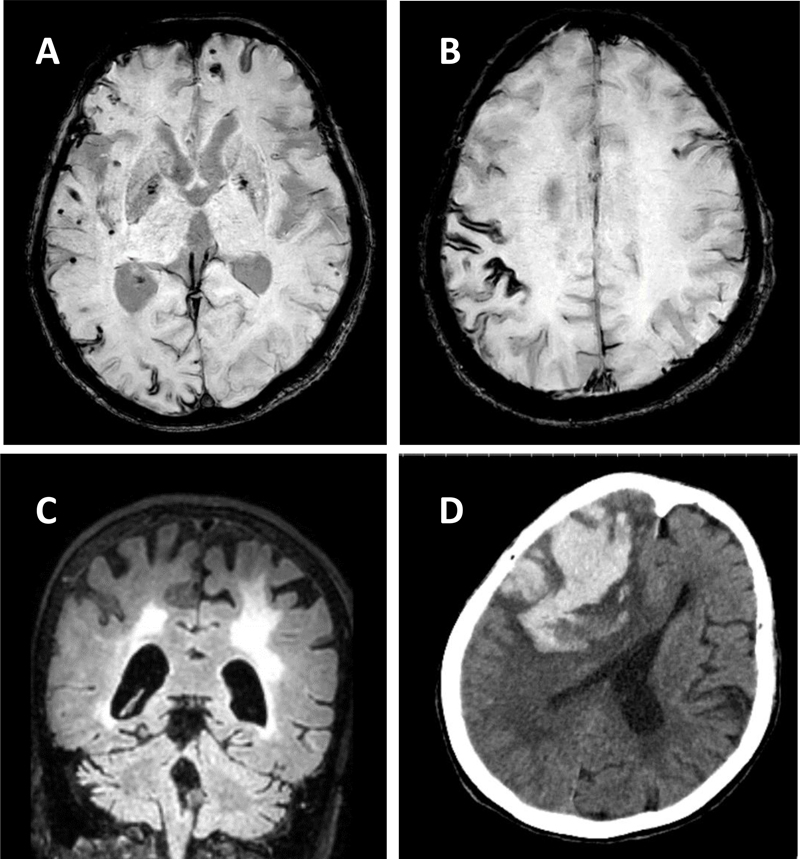
Figure 2. Embryonic neuronal cells from wild-type (WT) mice were implanted into the neocortex of pre-symptomatic 5xFAD transgenic mice. Aβ plaques developed as early as 4 weeks. Aβ can be transported by microglia and form plaques in the lesioned tissue. Microglia were involved in the propagation of Aβ pathology. Created with BioRender.com Topic 8: Perivascular space dilation is associated with vascular amyloid-β accumulation in the overlying cortex This work examined the enlargement of the perivascular space (PVS) in patients with cerebral amyloid angiopathy (CAA), an increasingly common form of cerebral small vessel disease. CAA is a common cause of lobar hemorrhage, cortical microhemorrhages and white matter disease in the elderly (Figure 3). In CAA, Aβ accumulates in the walls of cortical and leptomeningeal arteries, resulting in a loss of smooth muscle cells and impaired vascular function and can lead to lobar intracerebral hemorrhage in the elderly. The presence of cerebral vascular amyloid is a common neuropathological feature in patients with AD. PVS are compartments surrounding cerebral blood vessels that become visible on MRI when enlarged. It has been suggested that these spaces enlarge when there are deficits in dysfunctional perivascular clearance of waste products from the brain, including amyloid. The presence of enlarged PVS (EPVS) on MRI is associated with aging, high blood pressure, stroke and cognitive decline in other studies.
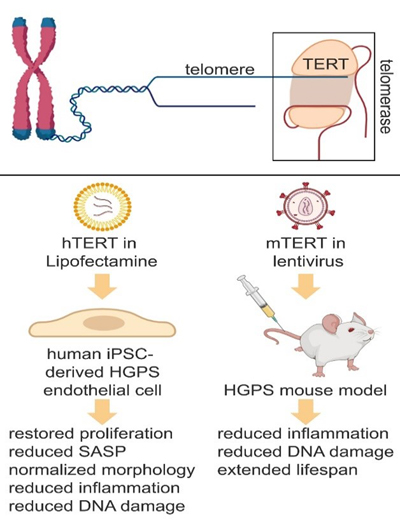
Figure 3. MRI and CT scan imaging of a 73-year-old woman with pathogenically confirmed cerebral amyloid angiopathy. Panel A shows innumerable cortical microhemorrhages, Panel B demonstrates cortical siderosis (GRE), and Panel C (FLAIR) shows extensive white matter disease and cerebral atrophy. Panel D is a CT scan taken 3 months after her prior MRI, when she presented with left hemiparesis and mental status changes. A large lobal hemorrhage is seen in the right frontal area, along with mass effect and cerebral edema. In this paper (10) the authors used 3T and 7T ex vivo MRI, semi-automatic segmentation and validated deep-learning-based models to quantify EPVS and associated histopathological abnormalities in 19 CAA cases and 5 controls. Severity of MRI-visible PVS during life was significantly associated with the severity of MRI-visible PVS on ex vivo MRI. This also corresponded with PVS enlargement on histopathology in the same areas. EPVS were located mainly around the white matter of perforating cortical arterioles. The amount of EPVS was associated with CAA severity in the overlying cortex. In addition, the authors found markedly reduced smooth muscle cells and increased vascular Aβ accumulation, extending into the WM, was seen in individual affected vessels with an EPVS. The presence of Aβ was also observed in the WM portion of the same perforating vessels, decreasing in intensity with increasing distance from the cortex. This could reflect blockage of Aβ clearance towards the brain surface, consistent with the decreasing severity of CAA between superficial and deep cortical layers, as previously shown histologically. These findings were confirmed in a patient with hereditary amyloid that was much younger (Dutch mutation). These findings are consistent with the concept that the development of EPVS reflects impaired outward flow along arterioles. This has implications for other small vessel diseases and suggests the importance of perivascular clearance mechanisms in human brain, which play an important role in the pathophysiology of both CAA and AD. These results support the concept of outward flow along the walls of cortical arterioles and suggest that impaired perivascular clearance of Aβ might lead to fluid stagnation and PVS enlargement around upstream portions of connected perivascular compartments. Interestingly, removal of Aβ plaques with anti-Aβ immunotherapy can worsen cortical and leptomeningeal CAA (11), suggesting that after plaque disaggregation Aβ is cleared along the cerebral vessels. Considering the currently unknown role of EPVS and white matter hyper-intensities (WMHs) and what they signify in humans both with and without AD/CAA, this paper has important clinical and pathophysiological relevance. Topic 9: Telomerase therapy reverses vascular senescence and extends lifespan in progeria mice Aging is a major risk factor for cardiovascular disease. With aging, the vasculature is altered, and molecular and structural changes occur in endothelial cells. Senescent endothelial cells show enhanced oxidative stress and DNA damages, aberrant chromatin changes, alteration in the nuclear envelope, telomere erosion, and increased production and secretion of pro-inflammatory molecules that negatively affect neighboring cells. Given that endothelial function regulates organ perfusion, inflammation and accommodates resident stem cells responsible for tissue repair, it is expected that restoring vascular homeostasis in aged organisms would improve organ function and extend lifespan. Hutchinson-Gilford progeria syndrome (HGPS) is a pediatric genetic disease characterized by accelerated aging. 90% of HGPS patients manifest cardiovascular disease early in life, in their teenage years. HGPS is caused by a mutation in a nuclear envelop protein (lamin A or progerin), which leads to deleterious consequences in nuclear morphology and gene expression. Unfortunately, treatments to mitigate this cardiovascular pathology are limited. The authors (11) used induced pluripotent stem cells (iPSCs) derived from HGPS patients, in addition to a HGPS mouse model to determine if reversing the senescence-associated phenotype in the vasculature had beneficial effects on lifespan and behavior in mice. They found that transient delivery of modified mRNA encoding for an enzyme that prevents telomere shortening (human telomerase, hTERT), which is a distinct feature of cell senescence, restored endothelial proliferation and function, upregulated lamin A, mitigated DNA damage, and reversed the inflammatory secretome seen in cultured HGPS endothelial cells (Figure 4). In addition, they delivered lentiviral mouse TERT (mTERT) to HGPS mice, and found that the aorta of treated mTERT mice showed reduced levels of VCAM-1 expression (an endothelial adhesion molecule that is increased with vascular inflammation), compared with control progeroid mice. These treated mice also exhibited reduced DNA damages in the endothelium of different tissues (aorta, lung, and liver), and their lifespan were significantly extended, compared with HGPS mice. This study demonstrates that TERT treatment restores senescence-associated phenotype in human HGPS endothelial cells, and expands life expectancy in a mouse model of accelerated aging. Its relevance lies in the importance of specifically targeting the endothelium to prevent vascular aging and mitigates cardiovascular disease associated with aging.
Figure 4. Telomerase (TERT) therapy reverses senescence-associated phenotypes in endothelial cells differentiated from Hutchinson-Gilford (HGPS) human iPSC lines and in a HGPS mouse model. Human TERT (hTERT) mRNA was delivered into hiPSC-derived (control and HGPS) endothelial cells by Lipofectamine transfection, and mouse TERT (mTERT) lentivirus construct was injected in 3 and 6 months old wild-type and HGPS mice via tail vein. Created with Biorender.com. Topic 10: Brain capillary pericytes exert a substantial but slow influence on blood flow The role of capillary pericytes, cells that line the capillary networks in the brain, on cerebral blood flow was investigated in this work from Dr. Shih’s group (12). The capillary bed has the highest flow resistance in the cerebral vasculature given their small diameter, and these vessels are lined with pericytes, unlike the larger arterioles of the brain in which neurovascular coupling is controlled largely by smooth muscle cells due to rapid blood flow demand and consist of α-SMA-positive mural cells. Two-photon microscopy and optogenetic stimulation was used to directly observe and manipulate brain capillary pericytes in vivo. To directly assess the contractile ability of capillary pericytes in vivo, the authors crossed PDGFRβ-Cre mice with reporter mice for the light-gated ion channel, ChR2-YFP. The authors confirmed that these cells, when stimulated optogenetically, can contract, decreasing their luminal diameter and reduce blood flow, but with slower kinetics than similar stimulation of mural cells on upstream pial and precapillary arterioles. Importantly, this vasoconstriction was associated with a decrease in red blood cell velocity and flux in the same capillaries, showing that the induced levels of constriction were sufficient to alter blood flow, and in fact led to significant “stalling” of RBC flow. Interestingly, “blebs” were seen in up to 30% of the pericytes. The authors hypothesized that these blebs reflected disruption of the actin cytoskeleton due to induced supra-physiological contraction. This slow vasoconstriction was inhibited by the clinically used vasodilator fasudil, a Rho-kinase inhibitor that blocks contractile machinery and promotes vasodilation. During optogenetic stimulation of capillary pericytes, fasudil attenuated relative and absolute capillary constriction in a dose-dependent manner and also led to a dose-dependent alleviation of persistent flow stalls. This has important translational relevance as this drug is used to treat cerebral vasospasm after subarachnoid hemorrhage in Japan, and could have a role in preventing the no reflow phenomenon in ischemic stroke after endovascular interventions. Capillary pericytes were also slower to constrict back to baseline following hypercapnia-induced dilation, and slower to dilate towards baseline following optogenetically induced vasoconstriction compared to pre-capillary arterioles. Optical ablation of single capillary pericytes led to sustained local dilation and a doubling of blood cell flux selectively in capillaries lacking pericyte contact. These data indicate that capillary pericytes contribute to basal blood flow resistance and slow modulation of blood flow throughout the brain. This could have major implications for other disorders related to cerebral hypoperfusion.
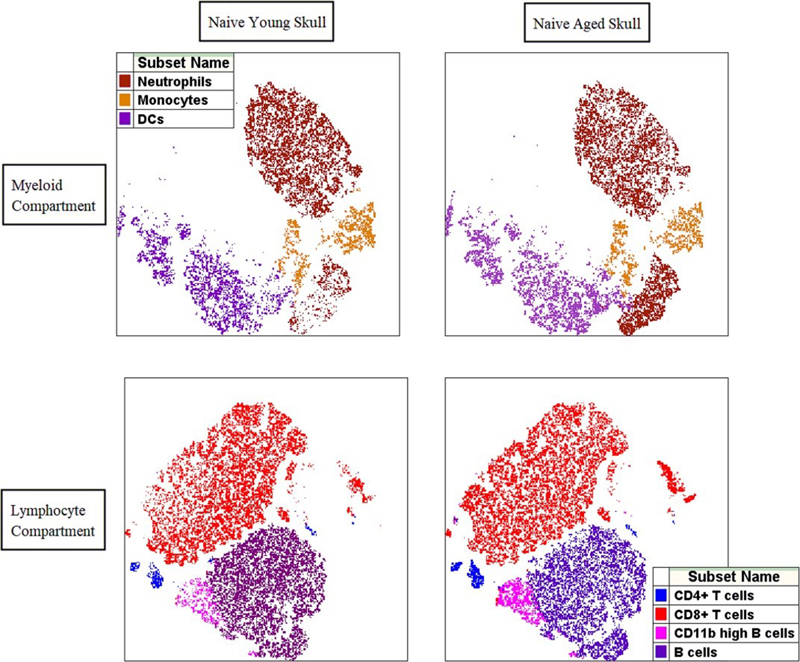
Figure 5. The skull bone marrow immune profile changes with aging. The composition of myeloid cell populations in the skull bone marrow changes with aging with significantly higher relative frequencies of neutrophils in aged skull (top). Skull bone marrow lymphocyte compartment contains significantly higher CD8+ T lymphocytes and activated CD11b high B lymphocytes in naïve aged mice when compared with young skull (bottom). Reprinted from Honarpisheh et al. (13). Conclusion Many exciting and novel papers were published in 2021. The compendium of papers highlighted demonstrate our growing understanding of vascular malformations, neuro-immune communication, and vascular function in aging and disease. One important caveat to consider is that many of these diseases predominately affect older individuals, such as stroke and amyloid related diseases. As the field moves forward it will be important to consider the age of the animals used, and also confirming that these pathways are involved in human disease using clinical samples to ensure translational relevance. For example, recent work in our own laboratory has shown that the skull immune compartment changes dramatically with age (Figure 5). Age-related changes in both peripheral and CNS immunity will be an important avenue of exploration in the future. Acknowledgements I would like to thank Felix Moruno Manchon, PhD, Juneyoung Lee, PhD, Pedram Honarpisheh, PhD, Aki Urayama, PhD, Sophie Ren, PhD, Alex Choi, MD, and Eunhee Kim, PhD, for their assistance in selection of these manuscripts. LDM is supported by the National Institute of Health (NIH) with grants from the NINDS (R37 NS096493, R01 NS108779) and NIA (R01 NS103592). References
Copyright: © 2022 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |