|
|
|
Free Neuropathology 3:13 (2022) |
|
Review |
|
Neurodegeneration: 2022 update |
|
John F. Crary |
|
Neuropathology Brain Bank & Research CoRE, Department of Pathology, Nash Family Department of Neuroscience, Department of Artificial Intelligence & Human Health, Ronald M. Loeb Center for Alzheimer's Disease, Friedman Brain Institute, Icahn School of Medicine at Mount Sinai, New York, NY, USA |
|
Corresponding author: |
|
Submitted: 18 April 2022 Accepted: 05 May 2022 Copyedited by: Georg Haase Published: 10 May 2022 |
|
Keywords: Neurodegeneration, Neuropathology, Aging, Microglia, Alzheimer disease, Tauopathy, Huntington’s disease, α-synucleinopathy, TDP-43 proteinopathy, Traumatic brain injury |
|
Abstract Here, we review a collection of recent manuscripts and research trends on the neuropathology of neurodegeneration that are considered by the author to be among the potentially most impactful. To the greatest extent possible, we chose to focus on histopathological studies that are most relevant to experimental and diagnostic neuropathology. While there has been an abundance of important recent discoveries and developments in neurodegenerative disease research, there was a deliberate effort here to provide balance to prevent disease categories and experimental approaches from overshadowing the others. The result is a diverse series of outstanding studies, together showing the landscape of progress across neurodegenerative disorders. One is a stereological study examining dystrophic microglia in aging. We highlight the first large genetic study of primary age-related tauopathy, showing convergence and divergence from classical Alzheimer’s disease. There were further advances in the neuropathological criteria and staging of chronic traumatic encephalopathy. Links suggesting a causal role for TMEM106B in TDP-43 proteinopathy emerged. Attempts to subtype Alzheimer’s disease on the molecular level were made. Evidence for a role for the VEGF family in cognitive impairment was advanced. Comparison of gene expression profiles from myeloid cells in peripheral blood and brain tissues from Parkinson’s disease patients revealed pathways that may lead to new mechanistic insights and biomarkers. A large autopsy series identified an increased frequency of central nervous system developmental malformations in Huntington’s disease. A robust and reliable system for assessing Lewy body pathology was proposed. Finally, we continue to be plagued by the COVID-19 pandemic, with lingering concerns of a long-term link with neurodegeneration. 1. Neurodegeneration-specific alterations in microglial morphology As the data from genetic, cellular and animal studies accumulates, the importance of microglial and neuroimmune function continues to grow. Microglia take on a number of different morphologies, including ramified, hypertrophic and dystrophic, that are demonstrable using various histopathological techniques. The importance of the dystrophic morphology is currently unclear and has been implicated in disease and age-related cellular senescence. Unfortunately, there has been a dearth of rigorous histopathological human studies looking at them across the aging spectrum. Critically, our understanding of dystrophic microglia has been hindered by a lack of well-designed and implemented quantitative studies of these various microglial morphological patterns across the human lifespan and in disease. Shahhidehpour et al. performed a stereological study, published in the Neurobiology of Aging, specifically investigating these microglial morphologies in human post-mortem brain tissues across the lifespan (Shahidehpour et al., 2021). The authors set out to test the hypothesis that age causes microglial dystrophy. Abnormalities in microglial function may be a strong contributor to age-related neurodegenerative disease, with cellular senescence being a critical hallmark of aging. The strength of the study was the rigorous stereological counts of microglial morphologies including hypertrophic and dystrophic types alongside total, across a collection of brains ranging in age in years from teenage to nineties (Figure 1). Surprisingly, while an age-associated increase in the total number of dystrophic microglia was found, this was not significant when compared to the total number of microglia which also increased with age. In contrast, the total number of dystrophic microglia was greater in subjects with a number of neurodegenerative diseases, including Alzheimer's disease, dementia with Lewy bodies, and limbic predominant age-related TDP-43 encephalopathy. The authors also provided evidence for a role of altered iron homeostasis in these dystrophic microglia using ferritin light chain as a marker of intracellular iron indicating a possible mechanism. This result pinpoints microglia dystrophy as a critical disease-associated signature in neurodegeneration.
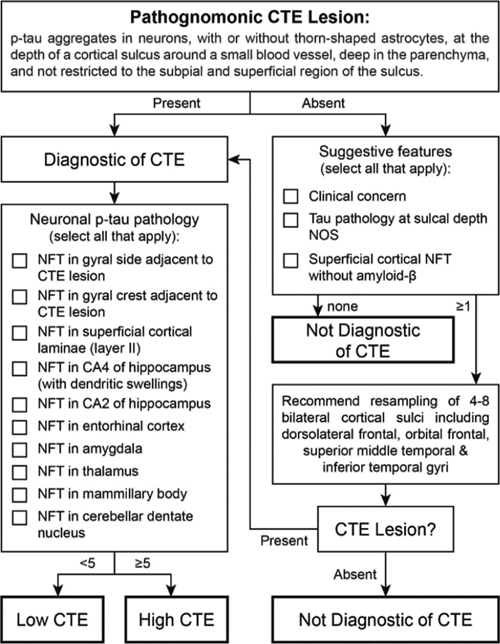
Figure 1. Examples of ramified (A, B), hypertrophic (C, D) and dystrophic microglia (E, F) reproduced with permission from Shahidehpour et al. (Shahidehpour et al., 2021). 2. Pathogenetic study in PART clarifies genetic overlap with AD and identifies JADE1 In 2014, a group of neuropathologists, led by the author of this article, defined criteria for a new category of neurodegenerative change, termed primary age-related tauopathy (PART) (Crary et al., 2014). Subsequently, there have been many clinical and neuropathological studies that have explored the contours and implications of this designation, both challenging and supporting its scientific validity as an independent age-associated neurodegenerative category. While the terminology has been broadly adopted, PART remains controversial and poorly understood. One of the biggest gaps in knowledge is the extent to which PART represents a distinct pathological process or shares mechanistic underpinnings with AD and perhaps the other tauopathies. This was addressed with the first genome-wide association study (GWAS) in primary age-related tauopathy published online in November 2021 in Acta Neuropathologica (Farrell et al., 2022). Given that PART is a ubiquitous neuropathological feature in essentially all aged human brains, Farrell et al. conducted a case-only quantitative trait analysis using Braak neurofibrillary stage as an endophenotype. While this was a relatively small GWAS (n=647), it was noted that most similar studies rely on clinical phenotypes, which are noisy relative to autopsy-derived endophenotypes, such as the Braak neurofibrillary tangle (NFT) stage used in this study. Looking at candidate genes implicated in Alzheimer’s disease and tauopathies revealed some associations, inducing with the MAPT H1 haplotype and other candidate loci, including SLC24A4, MS4A6A, HS3ST1 and EIF2AK3. As with previous studies, there was no association between PART and the most important and common risk allele for sporadic Alzheimer’s disease, APOE. The study also identified a novel locus on chromosome 4 that met genome-wide significance. Given that the locus contains a number of genes, the investigators used a computational approach, including examination of a single-cell tangle-containing RNA-seq dataset, to nominate gene for apoptosis and differentiation in epithelia 1 (JADE1) as the best candidate in the locus. The JADE1 protein has some interesting neuropathological features, co-localizing by immunohistochemistry with NFT in all tauopathies except Pick disease, the prototype tauopathy with three microtubule-binding domain repeats (3R). This was intriguing given that JADE1 protein biochemically interacts with only tau isoforms that contain 4 microtubule-binding domain repeats (4R). Finally, the JADE1 homolog in Drosophila, rhinoceros (rno), was shown to be a potent regulator of cell death in the fly eye and brain. Together, these findings suggest that JADE1 might be a component of a pathway that shared by many tauopathies. 3. Neuropathological criteria for chronic traumatic encephalopathy: consensus update As awareness and scientific interest in chronic traumatic encephalopathy (CTE) continues to expand, the neuropathological criteria continue to be a focus area. Concussions are extremely common, especially in military populations and contact sports athletes, and the long-term neurological sequelae remain under-investigated. Previously, the NIH convened a consensus panel of neuropathologists to establish preliminary consensus criteria for the neuropathological diagnosis of CTE (McKee et al., 2016). This landmark study has been highly influential, but questions persisted. For example, it was unclear as to what is the minimum requirement for a CTE diagnosis. Further, it was important to know whether CTE could be reliably differentiated from age-related tauopathies like primary age-related tauopathy (PART) and aging-related tau astrogliopathy (ARTAG). Further, a four-tiered staging system has been proposed but has not been independently validated (McKee et al., 2013). To address this, the NIH convened a second consensus panel meeting with eight neuropathologists expert in neurodegenerative tauopathies (Bieniek et al., 2021). The panel blindly reviewed 27 cases (17 CTE and 10 other tauopathies), applying the 2016 CTE criteria. First, there was substantial agreement in the diagnosis across raters which validated the robustness of the approach. Next, the panel turned to staging and developed a simplified and practical tool for the assessment of the CTE stage (Figure 2). The tool involves assessment of the 11 brain regions proposed by the first consensus meeting with a minimum threshold as a single pathognomonic lesion (with or without glial tangles). A point system was devised encompassing the numerous features of CTE with a cutoff of five, dichotomizing “low" and “high" CTE neuropathologic change. This simplified system had excellent correlation with the CTE stage as previously described (McKee et al., 2013). Numerous challenges and controversies in the field remain, but this represents a milestone in establishing rigor and reproducibility in CTE autopsy studies and provides a foundation for further advances in the field.
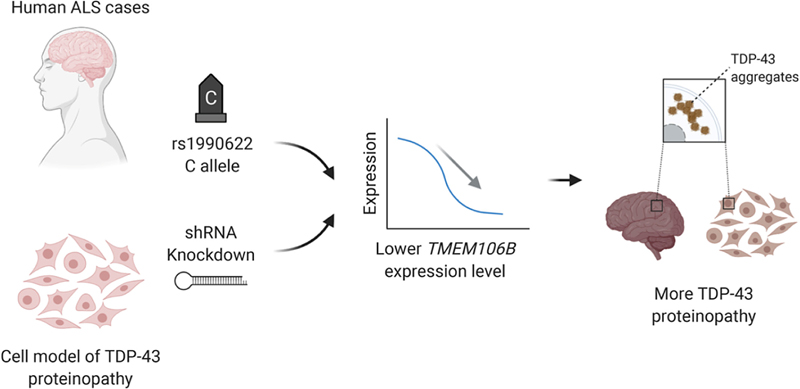
Figure 2. Flowchart for diagnosis of CTE reproduced with permission from Bieniek et al. (Bieniek et al., 2021). 4. A causal role for TMEM106B in TDP-43 pathology Inclusions containing the TAR DNA-binding protein 43 (TDP-43) are the hallmark neuropathological feature of a number of neurodegenerative diseases with divergent neuroanatomical vulnerability and symptomatology including amyotrophic lateral sclerosis (ALS), frontotemporal lobar degeneration (FTLD-TDP), and limbic age-related TDP-43 encephalopathy (LATE). The underlying reasons for these divergent pathoanatomical signatures are unclear and numerous factors likely contribute. While mutations in the TDP-43 gene are only rarely associated with TDP-43 proteinopathy, genetic variation in other genes including TMEM106B and C9orf72 are much more frequent in this context. To explore this, Mao et al. performed a rigorous pathoanatomical study published in Acta Neuropathologica examining the influence of genetic variation in TMEM106B and C9orf72 on the burden and distribution of TDP-43 proteinopathy in 899 brains from the University of Pennsylvania Center for Neurodegenerative Disease Research (CNDR) brain bank, including cases with ALS, FTLD-TDP, Alzheimer’s disease and Lewy body disease (Mao et al., 2021). TMEM106B risk alleles for FTLD-TDP were demonstrated to modify TDP-43 burden in ALS (but curiously not in FTLD-TDP). C9orf72 expansion was associated with TDP-43 burden in both ALS and FTLD-TDP. The interaction between TDP-43 and TMEM106B was confirmed in a cellular model. These data are intriguing because they suggest a causal role for TMEM106B in some forms of TDP-43 pathology (Figure 3).
Figure 3. Schematic showing the effects of TMEM106B on TDP-43 proteinopathy reproduced with permission from Mao et al. (Mao et al., 2021). 5. Defining molecular subtypes for Alzheimer’s disease According to a widely held dogma, Alzheimer’s disease arises secondarily from amyloid-beta toxicity that in turn leads to neurofibrillary degeneration and cell death. However, there is increasing evidence that AD is more heterogeneous than recognized, manifesting not only in the AD mimics such as limbic age-related TDP-43 encephalopathy and comorbid dementias such as Lewy body disease and cerebrovascular disease) but also within the group of subjects that meet the neuropathological criteria for AD. Neuropathological subtypes of AD have been proposed (Janocko et al., 2012; Murray et al., 2011) and gene expression studies in AD have been previously performed, but defining a molecularly defined subtype has never been attempted. Armed with a large RNA-seq dataset of over 1543 transcriptomes derived from four brain regions from two independent brain bank series, Neff et al. set out to define molecular subtypes of AD (Neff et al., 2021). To accomplish this, they leveraged a novel clustering method which uncovered five stable subtypes in three classes: A, B1/2 and C1/2 (Figure 4). One of these subtypes termed class C (“typical") which represented about one third of all cases, displayed expression changes similar to those previously reported in Alzheimer’s disease (Blalock et al., 2011) with increased immune response and decreased synaptic signaling. The class C cases also showed strong up-regulation of pathways involving amyloid-beta clearance, fiber formation and scavenger receptor activity. In contrast, expression changes in class A (“atypical") were generally opposite to class C (Blalock) changes. Class B (“intermediate") did not show changes in either direction. Tau-related pathways were strongly upregulated in both A and B1/2 subtypes. Certain pathway changes were subtype-specific illustrated by the up-regulation of protein degradation-related genes (e.g., ubiquitination) in class A or the up-regulation of organic acid-related genes in class B. Of note, the different classes were associated with distinct clinical and neuropathological phenotypes. Class C1 for instance showed a more frequent association with increased amyloid plaque burden than the tau-related class A and also had an increased APOE ε4 allele frequency whereas class A displayed increased MAPT pathway activation and decreased NFT association, thus potentially representing a resilient subtype. Many questions remain, especially how these subtypes align with previously proposed AD subtypes. Nevertheless, this study provides evidence that AD is likely not a single entity and paves the way towards additional studies addressing this critical barrier.
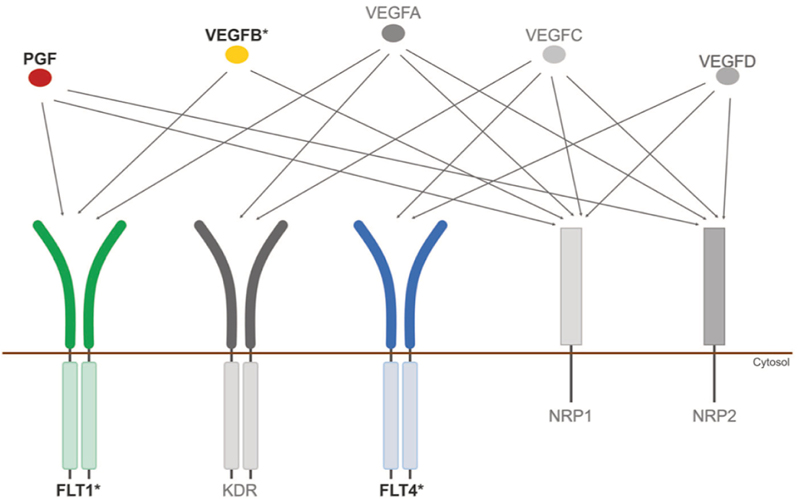
Figure 4. Molecular subtypes of Alzheimer’s disease defined by gene expression changes. Changes in the mean expression of gene pathways are indicated for AD subtypes A, B1, B2, C1 and C2 in comparison to normal control samples. Data from a previous study MSigDB are indicated on the right. Changes in gene expression are grouped by major areas of biological activity. Reproduced with permission from Neff et al. (Neff et al., 2021). 6. Cerebrovascular disease: VEGF gene family expression in cognitive aging and Alzheimer’s neuropathology Vascular endothelial growth factor (VEGF) is an angiogenic factor induced by hypoxia that plays an important role in angiogenesis. The VEGF gene family has several members which interact with a complex set of receptors and co-receptors which trigger (Figure 5) various signaling pathways highly relevant to neurodegeneration such as neurogenesis, neuroprotection, apoptosis, cell proliferation, and inflammation. Likewise, previous studies have linked VEGF signalling to various neurodegenerative and cerebrovascular diseases.
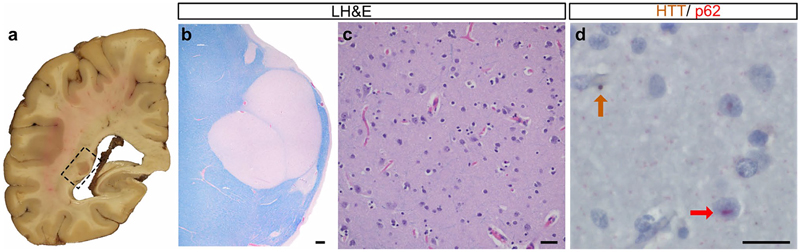
Figure 5. Illustration of the VEGF family of growth factors and their receptors. Asterisks indicate genes that are differentially expressed between AD and controls. Reproduced with permission from Mahoney et al. (Mahoney et al., 2021). A focused study published in Molecular Psychiatry by Mahoney et al. found intriguing direct evidence that changes in VEGF signaling are closely associated with the development of AD neuropathology and the progression of cognitive impairment (Mahoney et al., 2021). The authors leveraged the Religious Orders Study (ROS) and the Rush Memory and Aging Project (MAP), using global composited cognitive scores from 17 neuropsychological tests and autopsy measurements of amyloid plaque and tau burden from a total of 531 subjects with normal cognition, mild cognitive impairment (MCI) and AD-type dementia. The authors found that expression in the frontal cortex of four key components of VEGF signaling, VEGFB, FLT4, FLT1 and PGF, were associated with accelerated cognitive decline. All four of these genes were also associated with increased tau and amyloid burden. Additional work is required to elucidate exactly how VEGF signaling relates to AD neuropathology and cerebrovascular disease. This study, which requires additional confirmation and replication with molecular studies and cellular/animal modeling, spotlights the potential importance of VEGF in dementia. 7. Monocytes and microglia in Parkinson’s disease Evidence is accumulating that the immune system plays a critical role in Parkinson’s disease (PD). Investigating the pathogenic events occuring in critical immune cells especially in the PD brain remains of great interest. Myeloid cells and microglia have been understudied. Genetic studies have identified a large number of risk loci that alter the expression of nearby genes in easily accessible peripheral monocytes and there is enrichment of gene sets expressed in microglia. There are many functions of myeloid cells, including clearing of debris and maintaining brain homeostasis, that are highly relevant to PD pathogenesis. Further, alpha-synuclein can promote microglial release of neurotoxic factors potentially leading to death of dopaminergic neurons. Intriguingly, the possibility that alpha-synuclein pathology might be initiated in the enteric nervous system further highlights the potential importance of peripheral myeloid cells to influence pathogenesis. How myeloid cells are involved in PD is unclear and the extent to which specific immune cell types, in the periphery or in the central nervous system, participate remains a critical knowledge gap. In a paper published in Nature Aging by Navarro et al., an international team of researchers conducted a study looking at monocytes in peripheral blood as well as monocytes and microglia in post mortem brains from PD patients (Navarro et al., 2021). The authors sorted CD14+ peripheral monocytes and microglia from the blood and brains and performed a large-scale unbiased single cell transcriptomic study. The strength of this study are the direct comparisons between gene expression in the peripheral blood with the brain as most studies in PD have focused on dopaminergic neurons, fibroblasts, or whole blood. They identified genes and co-expression networks dysregulated in PD myeloid cells. They identified genes that are involved in mitochondrial and proteasomal function that are highly expressed in a proinflammatory intermediate subpopulation. They identified 17 variants that influence mRNA expression and PD risk. This enabled the authors to show that there are actually opposite mitochondrial transcriptomic profiles between microglia and macrophages (down regulated) versus peripheral monocytes (upregulated). These results provide further support for a prominent role for immune dysfunction in the pathogenesis of PD. 8. Neurodevelopmental alterations in Huntington’s disease Huntington’s disease (HD) is the prototype hyperkinetic movement disorder caused by polyQ repeat expansion in the huntingtin gene (HTT). While the neuropathology of HD has been extensively studied, accumulating evidence from cellular, animal and human studies pointed to the existence of neurodevelopmental abnormalities in HD. Of note, Barnat et al. have reported defects in neuronal migration in the brains of mutant HTT-fetuses and linked them to alterations in the interkinetic nuclear migration of progenitor cells (Barnat et al, Science 2020). Neuropathological evidence in adult human HD has however been lacking. In a paper published in Acta Neuropathologica, Hickman et al. provided the first neuropathological evidence that malformations are present in adult human HD (Hickman et al., 2021) through systematic evaluation of two large brain collections: a discovery cohort from the New York Brain Bank (NYBB) at Columbia University and a validation cohort from the Harvard Brain Tissue Resource Center (HBTRC), total n=3918. Of 130 HD brains examined from the NYBB, eight (6.2%) had at least one malformation compared to 12 (0.75%) in the 1600 non-HD brains. Periventricular nodular heterotopias (PNH) were the most frequent malformation (Figure 6), confirming abnormalities in neuronal migration in HD. Intriguingly, the neurons in the PNH contained p62-positive aggregates indicative of ongoing proteinopathy. Other cerebral malformations included hamartomas, cerebellar nodular heterotopias, and one multinodular vacuolating tumor of the cerebellum (MVNT). The frequency of HD-associated cerebral malformations was also significantly increased in the HBTRC cohort but at a lower frequency than in the NYBB cohort, which was likely explained by differences in sampling and assessment protocols. The asymmetric nature of the cerebral malformations in HD suggested that they were secondary to somatic mutation. Their higher frequency in women may be related to sexual dimorphism. The possibility that HD brains contain additional developmental or neurodevelopmental abnormalities merits further investigation. Nevertheless, this study provides concrete evidence that developmental abnormalities occur in human brains with HD and provide the foundation for a new avenue into studying HTT-related mechanisms.
Figure 6. A periventricular nodular heterotopia (PNH) in a 41-year-old woman with HD. Gross (a) and low power (LH&E stain) images (b) of a subependymal heterotopia containing non-dysmorphic neurons (c) that are immunopositive for HTT and p62 (d, arrows). Reproduced with permission from Hickman et al. (Hickman et al., 2021). 9. Criteria for Lewy body pathology: approaching consensus The spectrum of Lewy body disease (LBD) pathology includes Parkinson’s disease (PD), dementia with Lewy bodies (DLB) and PD with mild cognitive impairment or dementia. Currently, a neuropathological diagnosis of Lewy body disease can be rendered using any of four different proposed systems (Adler et al., 2019; Braak et al., 2003; Leverenz et al., 2008; McKeith et al., 2005). All use semi-quantitative assessment of the neuroanatomical distribution of Lewy pathology in neurites and cell bodies, but each has specific strengths and weaknesses. Each system suffers from issues related to interrater reliability and/or the inability to classify certain cases with Lewy pathology (LP), for example cases with olfactory bulb-only or amygdala-only disease. This stems from the heterogenous pathoanatomical distribution of LP. In 2021, new neuropathological consensus criteria that aim to address the limitations of these systems, termed the LP consensus criteria (LPC), were developed and published in Acta Neuropathologica in order to improve interrater reliability and to unequivocally classify all possible cases with LP (Attems et al., 2021). To improve inter-rater reliability, the authors implemented a dichotomized scoring system, with LP scored as either present or absent. The diagnostic categories were: olfactory-only, amygdala-predominant, brainstem, limbic, and neocortical LP. The 16 raters from 13 centers applied the LPC and each of the four existing systems to 34 cases with LP from the Newcastle Brain Tissue Resource and the University of Pennsylvania Brain Bank. Interrater reliability was good for the LPC, being comparable to the McKeith and Leverenz system (Krippendorff’s alpha ≈ 0.6), and considerably better than for Braak and Adler (Beach). The LPC system was as applicable as the Beach system and able to unequivocally classify essentially all cases, which was not the case for the Leverenz, McKeith and Braak systems which failed in 10%, 25% and nearly 30% of cases respectively. Given the reliability and applicability of the LPC system, we expect that it will be adopted broadly in the post-mortem evaluation of cases with Lewy body pathology. 10. Long-COVID intersecting with neurodegeneration During the COVID-19 pandemic, a number of neuropathological studies addressed the changes in the human brain from patients succumbing to acute SARS-CoV-2 infection. Microthrombi and strokes were among the most common findings together with indirect inflammatory and autoimmune changes but encephalitis was rare. After the peak of the pandemic, roughly one third of patients emerged with a clinical syndrom often termed post-acute sequelae of SARS-CoV-2 infection (PASC) and commonly referred to as “Long-COVID". Cardiac and pulmonary symptoms are prominent in PASC but there are also many alarming non-specific neurological and psychiatric symptoms including fatigue, cognitive complaints (“brain fog"), sensorimotor defects, headaches, insomnia, depression, and post-traumatic stress disorder (Taquet et al., 2021). A meta analysis confirmed decreased cognitive function seven months following COVID-19 infection (Crivelli et al., 2022). PASC has drawn attention in the field of neurodegeneration given the possibility that infection with the virus may trigger a long-term neurodegenerative process (Lennon, 2020). The mechanisms whereby PASC may be linked to neurodegeneration remained however elusive. Many investigators have raised the questions as to whether SARS-CoV-2 can cause neurodegeneration through its neurotropism or otherwise (Krey et al., 2021). Some investigators have proposed that SARS-CoV-2 enters the brain and may activate an AD-like program (Shen et al., 2022). The ACE2 COVID-19 receptor was indeed shown to be upregulated in the hippocampal formation of AD patients (Zhao et al., 2021), which may explain why medial temporal lobe structures might be vulnerable, potentially contributing to some post-infection memory impairment and neurodegeneration (Ritchie et al., 2020). Notably, signaling pathways leading to tau hyperphosphorylation in AD including GSK3β may be activated in post-mortem COVID-19 brains (Reiken et al., 2022). AD- and COVID-19 signaling pathways may converge at the level of the NRLP3 inflammasome through dysregulation of amyloid clearance by microglia (Cama et al., 2021, p. 3). Alternatively, SARS-CoV-2 infection may worsen AD for various reasons (Villa et al., 2022). While this constitutes perhaps the most urgent and potentially important research trend at this time, it remains to be seen whether there will be sustainable momentum. Acknowledgement Dr. Crary receives research funding from the NIH (R01AG054008, R01NS095252, R01AG062348, R01NS086736, U54NS115266, U54NS115322). We further acknowledge the Rainwater Charitable Trust/Tau Consortium, David and Elsie Werber, Alexander Saint Amand Scholar Award and Karen Strauss Cook Research Scholar Award. References Adler, C. H., Beach, T. G., Zhang, N., Shill, H. A., Driver-Dunckley, E., Caviness, J. N., Mehta, S. H., Sabbagh, M. N., Serrano, G. E., Sue, L. I., Belden, C. M., Powell, J., Jacobson, S. A., Zamrini, E., Shprecher, D., Davis, K. J., Dugger, B. N., & Hentz, J. G. (2019). Unified Staging System for Lewy Body Disorders: Clinicopathologic Correlations and Comparison to Braak Staging. Journal of Neuropathology and Experimental Neurology, 78(10), 891–899. https://doi.org/10.1093/jnen/nlz080 Attems, J., Toledo, J. B., Walker, L., Gelpi, E., Gentleman, S., Halliday, G., Hortobagyi, T., Jellinger, K., Kovacs, G. G., Lee, E. B., Love, S., McAleese, K. E., Nelson, P. T., Neumann, M., Parkkinen, L., Polvikoski, T., Sikorska, B., Smith, C., Grinberg, L. T., … McKeith, I. G. (2021). Neuropathological consensus criteria for the evaluation of Lewy pathology in post-mortem brains: A multi-centre study. Acta Neuropathologica, 141(2), 159–172. https://doi.org/10.1007/s00401-020-02255-2 Barnat, M., Capizzi, M., Aparicio, E., Boluda, S., Wennagel, D., Kacher, R., Kassem, R., Lenoir, S., Agasse, F., Braz, B.Y., Liu, J.-P., Ighil, J., Tessier, A., Zeitlin, S.O., Duyckaerts, C., Dommergues, M., Durr, A., Humbert, S., (2020). Huntington's disease alters human neurodevelopment. Science, 369(6505), 787-793. https://doi.org/10.1126/science.aax3338 Bieniek, K. F., Cairns, N. J., Crary, J. F., Dickson, D. W., Folkerth, R. D., Keene, C. D., Litvan, I., Perl, D. P., Stein, T. D., Vonsattel, J.-P., Stewart, W., Dams-O’Connor, K., Gordon, W. A., Tripodis, Y., Alvarez, V. E., Mez, J., Alosco, M. L., & McKee, A. C. (2021). The Second NINDS/NIBIB Consensus Meeting to Define Neuropathological Criteria for the Diagnosis of Chronic Traumatic Encephalopathy. Journal of Neuropathology and Experimental Neurology, 80(3), 210–219. https://doi.org/10.1093/jnen/nlab001 Blalock, E. M., Buechel, H. M., Popovic, J., Geddes, J. W., & Landfield, P. W. (2011). Microarray analyses of laser-captured hippocampus reveal distinct gray and white matter signatures associated with incipient Alzheimer’s disease. Journal of Chemical Neuroanatomy, 42(2), 118–126. https://doi.org/10.1016/j.jchemneu.2011.06.007 Braak, H., Del Tredici, K., Rüb, U., de Vos, R. A. I., Jansen Steur, E. N. H., & Braak, E. (2003). Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiology of Aging, 24(2), 197–211. https://doi.org/10.1016/s0197-4580(02)00065-9 Cama, V. F., Marín-Prida, J., Acosta-Rivero, N., Acosta, E. F., Díaz, L. O., Casadesús, A. V., Fernández-Marrero, B., Gilva-Rodríguez, N., Cremata-García, D., Cervantes-Llanos, M., Piniella-Matamoros, B., Sánchez, D., Del Rosario-Cruz, L., Borrajero, I., Díaz, A., González, Y., Pentón-Arias, E., Montero-González, T., Guillen-Nieto, G., & Pentón-Rol, G. (2021). The microglial NLRP3 inflammasome is involved in human SARS-CoV-2 cerebral pathogenicity: A report of three post-mortem cases. Journal of Neuroimmunology, 361, 577728. https://doi.org/10.1016/j.jneuroim.2021.577728 Crary, J. F., Trojanowski, J. Q., Schneider, J. A., Abisambra, J. F., Abner, E. L., Alafuzoff, I., Arnold, S. E., Attems, J., Beach, T. G., Bigio, E. H., Cairns, N. J., Dickson, D. W., Gearing, M., Grinberg, L. T., Hof, P. R., Hyman, B. T., Jellinger, K., Jicha, G. A., Kovacs, G. G., … Nelson, P. T. (2014). Primary age-related tauopathy (PART): A common pathology associated with human aging. Acta Neuropathologica, 128(6), 755–766. https://doi.org/10.1007/s00401-014-1349-0 Crivelli, L., Palmer, K., Calandri, I., Guekht, A., Beghi, E., Carroll, W., Frontera, J., García-Azorín, D., Westenberg, E., Winkler, A. S., Mangialasche, F., Allegri, R. F., & Kivipelto, M. (2022). Changes in cognitive functioning after COVID-19: A systematic review and meta-analysis. Alzheimer’s & Dementia: The Journal of the Alzheimer’s Association. https://doi.org/10.1002/alz.12644 Farrell, K., Kim, S., Han, N., Iida, M. A., Gonzalez, E. M., Otero-Garcia, M., Walker, J. M., Richardson, T. E., Renton, A. E., Andrews, S. J., Fulton-Howard, B., Humphrey, J., Vialle, R. A., Bowles, K. R., de Paiva Lopes, K., Whitney, K., Dangoor, D. K., Walsh, H., Marcora, E., … Crary, J. F. (2022). Genome-wide association study and functional validation implicates JADE1 in tauopathy. Acta Neuropathologica, 143(1), 33–53. https://doi.org/10.1007/s00401-021-02379-z Hickman, R. A., Faust, P. L., Rosenblum, M. K., Marder, K., Mehler, M. F., & Vonsattel, J. P. (2021). Developmental malformations in Huntington disease: Neuropathologic evidence of focal neuronal migration defects in a subset of adult brains. Acta Neuropathologica, 141(3), 399–413. https://doi.org/10.1007/s00401-021-02269-4 Janocko, N. J., Brodersen, K. A., Soto-Ortolaza, A. I., Ross, O. A., Liesinger, A. M., Duara, R., Graff-Radford, N. R., Dickson, D. W., & Murray, M. E. (2012). Neuropathologically defined subtypes of Alzheimer’s disease differ significantly from neurofibrillary tangle-predominant dementia. Acta Neuropathologica, 124(5), 681–692. https://doi.org/10.1007/s00401-012-1044-y Krey, L., Huber, M. K., Höglinger, G. U., & Wegner, F. (2021). Can SARS-CoV-2 Infection Lead to Neurodegeneration and Parkinson’s Disease? Brain Sciences, 11(12), 1654. https://doi.org/10.3390/brainsci11121654 Lennon, J. C. (2020). Neurologic and Immunologic Complications of COVID-19: Potential Long-Term Risk Factors for Alzheimer’s Disease. Journal of Alzheimer’s Disease Reports, 4(1), 217–221. https://doi.org/10.3233/ADR-200190 Leverenz, J. B., Hamilton, R., Tsuang, D. W., Schantz, A., Vavrek, D., Larson, E. B., Kukull, W. A., Lopez, O., Galasko, D., Masliah, E., Kaye, J., Woltjer, R., Clark, C., Trojanowski, J. Q., & Montine, T. J. (2008). Empiric refinement of the pathologic assessment of Lewy-related pathology in the dementia patient. Brain Pathology (Zurich, Switzerland), 18(2), 220–224. https://doi.org/10.1111/j.1750-3639.2007.00117.x Mahoney, E. R., Dumitrescu, L., Moore, A. M., Cambronero, F. E., De Jager, P. L., Koran, M. E. I., Petyuk, V. A., Robinson, R. A. S., Goyal, S., Schneider, J. A., Bennett, D. A., Jefferson, A. L., & Hohman, T. J. (2021). Brain expression of the vascular endothelial growth factor gene family in cognitive aging and alzheimer’s disease. Molecular Psychiatry, 26(3), 888–896. https://doi.org/10.1038/s41380-019-0458-5 Mao, F., Robinson, J. L., Unger, T., Posavi, M., Amado, D. A., Elman, L., Grossman, M., Wolk, D. A., Lee, E. B., Van Deerlin, V. M., Porta, S., Lee, V. M. Y., Trojanowski, J. Q., & Chen-Plotkin, A. S. (2021). TMEM106B modifies TDP-43 pathology in human ALS brain and cell-based models of TDP-43 proteinopathy. Acta Neuropathologica, 142(4), 629–642. https://doi.org/10.1007/s00401-021-02330-2 McKee, A. C., Cairns, N. J., Dickson, D. W., Folkerth, R. D., Keene, C. D., Litvan, I., Perl, D. P., Stein, T. D., Vonsattel, J.-P., Stewart, W., Tripodis, Y., Crary, J. F., Bieniek, K. F., Dams-O’Connor, K., Alvarez, V. E., Gordon, W. A., & TBI/CTE group. (2016). The first NINDS/NIBIB consensus meeting to define neuropathological criteria for the diagnosis of chronic traumatic encephalopathy. Acta Neuropathologica, 131(1), 75–86. https://doi.org/10.1007/s00401-015-1515-z McKee, A. C., Stern, R. A., Nowinski, C. J., Stein, T. D., Alvarez, V. E., Daneshvar, D. H., Lee, H.-S., Wojtowicz, S. M., Hall, G., Baugh, C. M., Riley, D. O., Kubilus, C. A., Cormier, K. A., Jacobs, M. A., Martin, B. R., Abraham, C. R., Ikezu, T., Reichard, R. R., Wolozin, B. L., … Cantu, R. C. (2013). The spectrum of disease in chronic traumatic encephalopathy. Brain: A Journal of Neurology, 136(Pt 1), 43–64. https://doi.org/10.1093/brain/aws307 McKeith, I. G., Dickson, D. W., Lowe, J., Emre, M., O’Brien, J. T., Feldman, H., Cummings, J., Duda, J. E., Lippa, C., Perry, E. K., Aarsland, D., Arai, H., Ballard, C. G., Boeve, B., Burn, D. J., Costa, D., Del Ser, T., Dubois, B., Galasko, D., … Consortium on DLB. (2005). Diagnosis and management of dementia with Lewy bodies: Third report of the DLB Consortium. Neurology, 65(12), 1863–1872. https://doi.org/10.1212/01.wnl.0000187889.17253.b1 Murray, M. E., Graff-Radford, N. R., Ross, O. A., Petersen, R. C., Duara, R., & Dickson, D. W. (2011). Neuropathologically defined subtypes of Alzheimer’s disease with distinct clinical characteristics: A retrospective study. The Lancet. Neurology, 10(9), 785–796. https://doi.org/10.1016/S1474-4422(11)70156-9 Navarro, E., Udine, E., de Paiva Lopes, K., Parks, M., Riboldi, G., Schilder, B. M., Humphrey, J., Snijders, G. J. L., Vialle, R. A., Zhuang, M., Sikder, T., Argyrou, C., Allan, A., Chao, M. J., Farrell, K., Henderson, B., Simon, S., Raymond, D., Elango, S., … Raj, T. (2021). Dysregulation of mitochondrial and proteolysosomal genes in Parkinson’s disease myeloid cells. Nature Aging, 1(9), 850–863. https://doi.org/10.1038/s43587-021-00110-x Neff, R. A., Wang, M., Vatansever, S., Guo, L., Ming, C., Wang, Q., Wang, E., Horgusluoglu-Moloch, E., Song, W.-M., Li, A., Castranio, E. L., Tcw, J., Ho, L., Goate, A., Fossati, V., Noggle, S., Gandy, S., Ehrlich, M. E., Katsel, P., … Zhang, B. (2021). Molecular subtyping of Alzheimer’s disease using RNA sequencing data reveals novel mechanisms and targets. Science Advances, 7(2), eabb5398. https://doi.org/10.1126/sciadv.abb5398 Reiken, S., Sittenfeld, L., Dridi, H., Liu, Y., Liu, X., & Marks, A. R. (2022). Alzheimer’s-like signaling in brains of COVID-19 patients. Alzheimer’s & Dementia: The Journal of the Alzheimer’s Association. https://doi.org/10.1002/alz.12558 Ritchie, K., Chan, D., & Watermeyer, T. (2020). The cognitive consequences of the COVID-19 epidemic: Collateral damage? Brain Communications, 2(2), fcaa069. https://doi.org/10.1093/braincomms/fcaa069 Shahidehpour, R. K., Higdon, R. E., Crawford, N. G., Neltner, J. H., Ighodaro, E. T., Patel, E., Price, D., Nelson, P. T., & Bachstetter, A. D. (2021). Dystrophic microglia are associated with neurodegenerative disease and not healthy aging in the human brain. Neurobiology of Aging, 99, 19–27. https://doi.org/10.1016/j.neurobiolaging.2020.12.003 Shen, W.-B., Logue, J., Yang, P., Baracco, L., Elahi, M., Reece, E. A., Wang, B., Li, L., Blanchard, T. G., Han, Z., Frieman, M. B., Rissman, R. A., & Yang, P. (2022). SARS-CoV-2 invades cognitive centers of the brain and induces Alzheimer’s-like neuropathology. BioRxiv: The Preprint Server for Biology, 2022.01.31.478476. https://doi.org/10.1101/2022.01.31.478476 Taquet, M., Geddes, J. R., Husain, M., Luciano, S., & Harrison, P. J. (2021). 6-month neurological and psychiatric outcomes in 236 379 survivors of COVID-19: A retrospective cohort study using electronic health records. The Lancet. Psychiatry, 8(5), 416–427. https://doi.org/10.1016/S2215-0366(21)00084-5 Villa, C., Rivellini, E., Lavitrano, M., & Combi, R. (2022). Can SARS-CoV-2 Infection Exacerbate Alzheimer’s Disease? An Overview of Shared Risk Factors and Pathogenetic Mechanisms. Journal of Personalized Medicine, 12(1), 29. https://doi.org/10.3390/jpm12010029 Zhao, Y., Li, W., & Lukiw, W. (2021). Ubiquity of the SARS-CoV-2 receptor ACE2 and upregulation in limbic regions of Alzheimer’s disease brain. Folia Neuropathologica, 59(3), 232–238. https://doi.org/10.5114/fn.2021.109495
Copyright: © 2022 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |