|
|
|
Free Neuropathology 2:21 (2021) |
|
Original Paper |
|
Differential gene expression in the cortical sulcus compared to the gyral crest within the early stages of chronic traumatic encephalopathy |
|
Jonathan D. Cherry*1,2,3,4, Filisia Agus2,5, Erin Dixon3,4, Bertrand Huber2,3,4,6, Victor E. Alvarez2,3,4,7, Jesse Mez2,3, Ann C. McKee1,2,3,4,7, Adam Labadorf2,5,6, Thor D. Stein1,3,4,7 |
|
1 Department of Pathology and Laboratory Medicine, Boston University School of Medicine, Boston MA, USA |
|
* Corresponding authors: |
|
Submitted: 21 July 2021 Accepted: 12 August 2021 Copyedited by: Lauren Walker Published: 17 August 2021 |
|
Additional resources and electronic supplementary material: download supplementary material |
|
Keywords: CTE, RNA-seq, Repetitive head trauma, Inflammation, Tau, TBI |
|
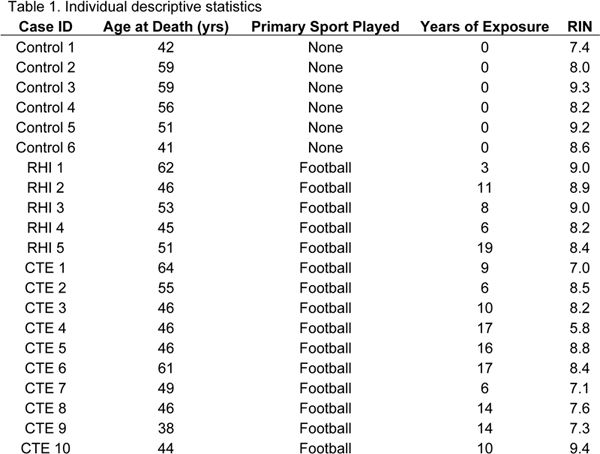
Abstract Chronic traumatic encephalopathy (CTE) is a progressive neurodegenerative tauopathy found in individuals with a history of repetitive head impacts (RHI). Previous work has demonstrated that neuroinflammation is involved in CTE pathogenesis, however, the specific inflammatory mechanisms are still unclear. Here, using RNA-sequencing and gene set enrichment analysis (GSEA), we investigated the genetic changes found in tissue taken from the region CTE pathology is first found, the cortical sulcus, and compared it to neighboring gryal crest tissue to identify what pathways were directly related to initial hyperphosphorylated tau (p-tau) deposition. 21 cases were chosen for analysis: 6 cases had no exposure to RHI or presence of neurodegenerative disease (Control), 5 cases had exposure to RHI but no presence of neurodegenerative disease (RHI), and 10 cases had exposure to RHI and low stage CTE (CTE). Two sets of genes were identified: genes that changed in both the sulcus and crest and genes that changed specifically in the sulcus relative to the crest. When examining genes that changed in both the sulcus and crest, GSEA demonstrated an increase in immune related processes and a decrease in neuronal processes in RHI and CTE groups. Sulcal specific alterations were observed to be driven by three mechanisms: anatomy, RHI, or p-tau. First, we observed consistent sulcal specific alterations in immune, extracellular matrix, vascular, neuronal, and endocytosis/exocytosis categories across all groups, suggesting the sulcus has a unique molecular signature compared to the neighboring crest independent of pathology. Second, individuals with a history of RHI demonstrated impairment in metabolic and mitochondrial related processes. Finally, in individuals with CTE, we observed impairment of immune and phagocytic related processes. Overall, this work provides the first observation of biological processes specifically altered in the sulcus that could be directly implicated in CTE pathogenesis and provide novel targets for biomarkers and therapies. Introduction Chronic traumatic encephalopathy (CTE) is a progressive neurodegenerative disease found in individuals with a history of exposure to repetitive head impacts (RHI) typically received through playing contact sports such as American football, hockey, soccer, boxing, or rugby, or through military service-related injuries like blasts [3, 23, 24]. Currently, CTE can only be definitively diagnosed after death through neuropathologic autopsy. The pathognomonic lesion that defines the disease consists of perivascular hyperphosphorylated tau (p-tau) found in neurons and sometimes astrocytes, initially at the depths of the cortical sulcus in the frontal cortex [3]. Through computational modeling and helmet sensor data, it was observed that the sulcus and blood vessels receive the greatest amount of damage after head trauma, directly linking RHI induced trauma to CTE pathogenesis [11]. However, the specific biological response after trauma that drives CTE pathology is still unclear. Under normal circumstances, inflammation is a necessary part of the injury cascade. In the brain, neuroinflammation has been observed to be necessary for wound healing responses after damage, triggering an innate and adaptive immune response to fight infectious agents, and recruit beneficial immune cells to areas of need [6]. However, prolonged or chronic inflammation can be harmful and can result in tissue damage and long-term pathology. RHI has been shown to induce a chronic neuroinflammatory environment in both mouse and human studies [9, 10, 32]. Additionally, previous work from our laboratory has demonstrated that elevated microglial recruitment via the chemokine CCL2, and glial activation was related to greater years playing contact sports and is a prominent feature in CTE [4, 7]. We observed that microglia-mediated neuroinflammation may exist in a feedback loop with p-tau pathology. Neuroinflammation induces p-tau deposition, which in turn induces more neuroinflammation leading to a vicious cycle, similar to the inflammatory cascade hypothesis in Alzheimer’s disease (AD) [12, 16]. However, the concept of neuroinflammation is broad, encompassing inflammatory, anti-inflammatory, senescent, and immunoregulatory features that previous histology-based studies were unable to easily segregate [6]. Therefore, there is a critical need to understand the individual inflammatory phenotypes that emerge during RHI to have more complete insight into the various mechanisms which may directly result in CTE pathogenesis. To help fill in knowledge gaps, high level bioinformatic analyses are needed. Previous studies have demonstrated that high throughput genomic techniques and bioinformatic pipelines are an optimal method to investigate large scale changes in the brain and determine specific pathways and effects in a more efficient manner [1, 17]. Herein, we utilized bulk tissue RNA-sequencing targeted to cortical anatomy, in order to investigate the spectrum of inflammatory and neurodegenerative phenotypes that occur as a result of RHI and during early stage CTE. As the earliest neuropathologic feature of CTE is p-tau pathology found at the depth of the cortical sulcus, we chose to examine what genetic changes occur specifically in the sulcus compared to the neighboring gyral crest. By directly comparing between the sulcus and crest within each case, we will have unique insights into which genes or biological processes are specifically altered in the sulcus as opposed to genes that are part of a more tissue wide response. We will utilize three separate groups, individuals without any RHI or CTE (controls), individuals with RHI but no CTE (RHI), and individuals with early stage CTE (CTE). Comparison of these three groups will allow direct analysis of different stages of disease. Furthermore, the results will provide new understandings and insight into what genes and biological processes might be directly related to initial CTE p-tau deposition as opposed to a general CNS response to trauma. Finally, genes that are found to be specific to CTE related pathology will become targets for future biomarker or therapeutic intervention studies. Methods Subjects A convenience sample of 15 brain donors with a history of RHI exposure from contact sports were selected from the Understanding Neurological Injury and Traumatic Encephalopathy (UNITE) brain bank. Selection was based on availability of frozen frontal cortex tissue and if the cases were male and between the ages of 35-65 years old. Samples were excluded from the study if they carried a neuropathologic diagnosis of AD, neocortical Lewy bodies, frontotemporal lobar degeneration (FTLD), or motor neuron disease (MND) based on established neuropathologic criteria for each disease [19, 20, 25, 26]. An additional 6 donors were obtained from the National Post-Traumatic Stress Disorder (PTSD) brain bank (National Center for PTSD, VA Boston Healthcare System, Boston MA). These 6 donors were selected from a group that lacked a history of RHI, did not have evidence of a neurodegenerative disease at autopsy, and were not diagnosed with PTSD in life. Samples were grouped into three categories: 6 cases with no history of repetitive head trauma or neurodegenerative disease (Control), 5 cases with a history of repetitive head trauma but no neurodegenerative disease (RHI), and 10 cases with a history of repetitive head trauma and a diagnosis of low stage CTE (CTE). There was no difference in the mean age of death of each group: Control- 51.3 years old ± 8.2, RHI- 51.4 years old ± 6.8, CTE- 49.5 years old ± 8.1. All cases were male and had a history of playing American football. Individual descriptive statistics for the age at death, years of exposure, sport played, and RNA integrity number (RIN), are provided in Table 1.
Next-of-kin provided written consent for participation and donation. Institutional review board approval for brain donation was obtained through the Boston University Alzheimer’s Disease and CTE center, Human Subjects Institutional Review Board of the Boston University School of Medicine, and Edith Nourse Rogers Memorial Veterans Hospital (Bedford, MA). Neuropathological Assessment Post-mortem fresh frozen tissue from the dorsolateral frontal cortex was obtained using previously described procedures [4]. Neuropathologic diagnosis was obtained from formalin fixed paraffin embedded tissue as previously described [21, 24]. Briefly, 22 sections of paraffin embedded tissue were stained for hyperphosphorylated tau (p-tau), alpha-synuclein, amyloid beta, TDP-43, luxol fast blue, and hematoxylin and eosin using previously described methods [22]. A neuropathologic diagnosis of CTE was made using National Institute of Neurological Disorders and Stroke (NINDS) consensus criteria [3, 21]. CTE cases could be subdivided based on the presence, extent, and severity of p-tau deposition through the brain. As the focus of the study was to examine early changes, only cases that met the “low stage CTE” criteria were used for analysis. The low stage CTE designation encompasses CTE stage I&II [2, 21] and shows strong agreement with the recent NINDS consensus staging for low stage CTE [3]. Neuropathologic examination occurred blinded to clinical results. All evaluations were reviewed by four neuropathologists (VA, BH, TS, AM); discrepancies in the diagnosis were resolved by consensus conference. Demographics, athletic history (type of sports played, level, position, age of first exposure to sports and years playing contact sports), military history (branch, location of service and duration of combat exposure), and traumatic brain injury (TBI) history (including number of concussions) were queried during a telephone interview as detailed previously [30]. RNA-Sequencing 50µg of fresh frozen tissue was taken from the dorsolateral frontal cortex grey matter at two locations for each case: the depth of the superior frontal cortical sulcus (defined as the bottom third of the sulcus) and the associated neighboring gryal crest at the level of the anterior caudate nucleus. Frozen tissue was process using a mortar and pestle, and mRNA was extracted and isolated using a Maxwell RNA extraction kit (Promega) as per manufactures instruction. An Ion Apliseq Transcriptome Human Gene Expression Kit (ThermoFisher Scientific) was then used to convert mRNA to cDNA and establish human transcriptome cDNA libraries as per manufactures instructions. Human transcriptome libraries were then sequenced using an Ion Torrent S5 next generation sequencer (ThermoFisher Scientific). Samples were processed and run across two batches. Batch one contained 6 control and 6 CTE cases, and batch two contained 5 RHI and 4 CTE cases and were compared separately to avoid batch effects. Normalized counts for each sample were determined using the Ion Torrent Suit Software 5.10 and concatenated together into genes by sample count matrix. The normalized counts were then transformed with Rlog for principal component analysis (PCA). Differential Expression analysis Genes with more than 50% zero counts within each group were filtered out. Pairwise differential expression (DE) analyses were conducted separately for CTE vs RHI and CTE vs Control using DESeq2 bioconductor package, modeling counts as a function of case status adjusting for age at death [18]. To analyze changes between the sulcus and crest within each group, a ratio between the sulcus and crest genes were calculated using the concatenated normalized counts matrix. Genes that could not be detected in all samples (genes with 0 counts for all samples) were filtered out. A pseudocount of 0.01 was added to the counts to prevent infinity values from zero divisions. The ratio was then calculated as log 2 of sulcus counts divided by crest counts for each gene within each sample. To attenuate extreme ratios due to small count values in either tissue, the ratios were arcsin transformed by first dividing by 0.5 and then applying the arcsin function using the NumPy python package. This procedure results in a log2 fold change of sulcus vs crest for each gene in each sample. To identify genes that exhibited significantly different expression between tissues within each case status, one-sample, two-tailed t-tests comparing with an expected mean log2 fold change of zero were performed for the fold changes of each gene. p-values were adjusted for multiple hypotheses using the Benjamini-Hochberg procedure. Similarly, two-sample t-tests were run to compare the sulcus vs crest fold changes for CTE vs RHI, CTE vs Control, and RHI vs Control. Gene set enrichment analysis (GSEA) Gene set enrichment analysis for all DE gene lists were performed using the fgsea R package in Bioconductor and Gene Ontology (GO) geneset annotations from GOAtools python package [14, 29]. GSEA statistics were calculated using gene list sorted by descending log2 fold change (from DE) or log 2 ratio t-statistics, and significance was assessed for gene sets at FDR < 0.1. The resulting normalized enrichment scores (NES) were then used to create heatmaps. GSEA biological process were grouped into 1 out of 11 biological categories by hand, based on expert opinion. Categories were discussed and confirmed among 3 authors (JC, AL, TS). Any differences were resolved through consensus conference. The biological categories were: immune, vascular, neuronal, extracellular matrix (ECM), mitochondrial/metabolism, signaling, cytoskeleton, transcription/translation, endocytosis/exocytosis, protein processing, and any process that didn’t fit the previous 10 category was assigned to “other”. Finally, gene ontology analysis from common or unique genes groups was carried out using Metascape [33]. All code and supplementary information needed to reproduce this analysis is available at https://osf.io/exsfa/. Results Overall, 3 differential expression analyses were carried out. A summary of each comparison is outlined in Figure 1. The first analysis was to directly compare the differentially expressed genes in the sulcus or crest to the matching region across control, RHI, or CTE groups (Figure 1A, Comparison 1). The second analysis was to directly compare the sulcus vs. crest within each group (Figure 1B, Comparison 2). The final analysis was to compare how the sulcus vs. crest relative ratios of each gene changed across each disease group (Figure 1C, Comparison 3).
Figure 1. Summary of differential expression analysis used for comparisonsRepresentative images demonstrating how which regions were being used for comparison. A) Comparison 1 compares the differentially expressed genes in the sulcus and crest to the corresponding region between control, RHI, and CTE. B) Comparison 2 directly compares the sulcus to the crest within each group. C) Comparison 3 uses the ratio created from comparison 2 and compares that sulcus vs crest ratio from RHI and CTE groups to the control group to determine how the sulcal specific genes changes over the course of disease. Differentially expressed genes had similar directions of effect in the sulcus and crest We first examined the differentially expressed (DE) genes found in the sulcus and crest to identify the genes most affected in early CTE compared with control and RHI groups (Table 2) (Figure 1A, Comparison 1).
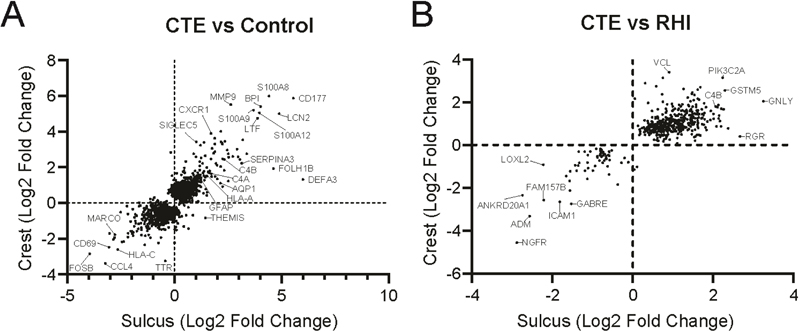
The relative sulcus and crest DE genes that were significantly altered (nominal p < 0.05) in at least one region were plotted against each other to examine if there were similar direction changes occurring in the sulcus and crest (Figure 2). Changes that were consistent across both sulcus and crest could be interpreted as a general frontal cortex wide response. First, we compared CTE to Control (Figure 2A). We observed that 95% of genes had the same direction of effect in both the sulcus and crest. Only 109 (5.0% total DE genes) genes had an opposite direction of effect (Figure 2A). When examining the changes between CTE and RHI, we observed fewer genes were altered compared to CTE vs Control (Table 2). Almost all of the genes had the same direction of effect (Figure 2B). Only 2 genes (0.4% total DE genes), JMJD6 and VAT1L, were elevated in the sulcus but decreased in the crest. The genes with the largest change, in addition to specific genes of interest were highlighted. A full list of DE genes for the CTE vs Control and CTE vs RHI analysis can be found in Supplementary file 1.
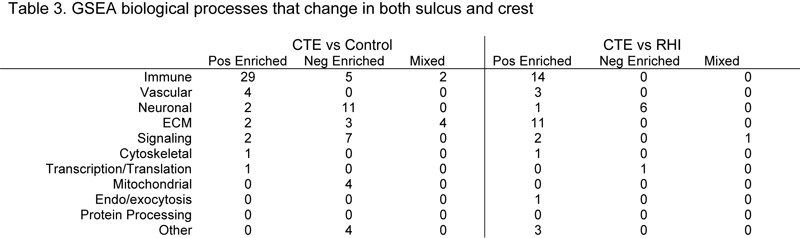
Figure 2. Comparative analysis of the differentially expressed genes found in the sulcus and crestAnalysis of DE genes from the sulcus and crest plotted against each other comparing (A) CTE vs Control and (B) CTE vs RHI. Genes were plotted if they were significantly altered (p<0.05) in at least one region. Each dot represents one gene. Genes of interest and the top expressed have been annotated. Gene set enrichment analysis (GSEA) was then performed on each set of genes to characterize biological processes that are altered in CTE (Table 3). Each GSEA process was then assigned to 1 of 11 overall biological categories to better contextualize changes (Figure 3). When comparing CTE vs Control (Figure 3A), 81 processes were observed to have an FDR adjusted p < 0.1 in the sulcus or the crest. 41 processes were positively enriched and 34 were negatively enriched in the same direction in both the sulcus and crest. A complete breakdown of group assignments can be found in Table 3. The majority of positively enriched processes were involved the immune category with 70.7% of total processes. Conversely, only 14.7% of the negatively enriched processes were immune processes. Neuronal processes were the largest group of negatively enriched processes with 35.5%. There were fewer positively enriched neuronal processes (4.9%). Most signaling processes were negatively enriched (20.6%) compared to positively enriched (4.9%). Mitochondria processes were only found to be negatively enriched, while vascular processes were only found in the positively enriched group (Table 3). 6 processes were found to have a mixed direction of effect. Of those 6 processes, 2 were immune and 4 were extra-cellular matrix (ECM) related (Table 3).
For CTE vs RHI (Figure 3B), 44 processes were observed to have an FDR adjusted p < 0.1 in the sulcus or crest. 36 processes were positively enriched, and 7 processes were negatively enriched in the same direction. The largest positively enriched category was immune processes with 38.9%. No immune processes were negatively enriched. ECM processes were the second largest positively enriched category with 30.6% of processes. No ECM processes were negatively enriched. Neuronal processes were the majority of negatively enriched processes (85.7%) and were higher than positively enriched neuronal processes. The only other negatively enriched process was transcription/translation (14.3%). Vascular, signaling, cytoskeletal, endo/exocytosis, and other processes were all similarly expressed. One process, “signaling receptor activity”, from the signaling category, was decreased in the crest and increased in the sulcus (Figure 3).
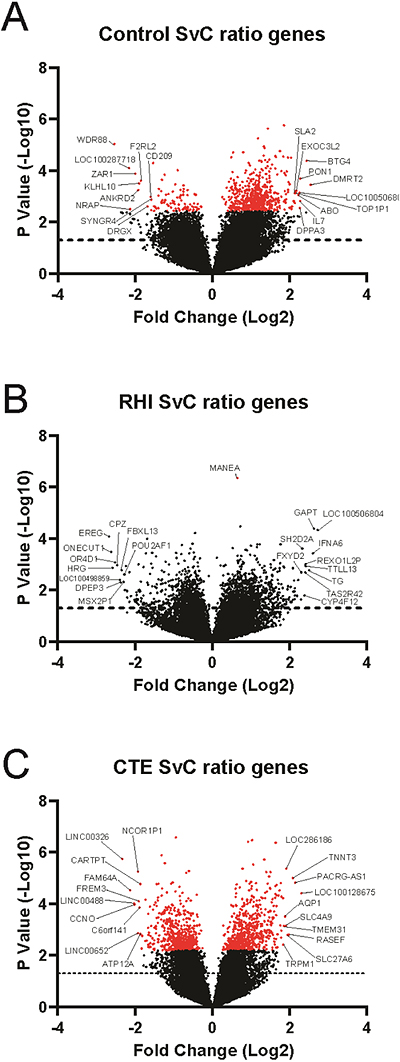
Figure 3. Heatmap of sulcus and crest GSEA biological processesUsing the DE genes from the sulcus and crest, GSEA biological processes were derived and plotted in a heatmap comparing (A) CTE vs Control and (B) CTE vs RHI. Processes were included if they were significantly enriched (FDR adjusted p < 0.1) in at least one region. Each GSEA biological process was assigned to 1 of 11 biological categories to better identify overall changes. [Please click on the figure to download a high-resolution version] There were significant differences in the sulcus compared to the crest We next chose to directly compare the sulcus and crest within the controls, RHI, and CTE groups to determine which genes were specifically altered in the sulcus compared with the crest by computing an arcsin transformed log2 ratio of sulcus vs crest normalized counts within individuals (see Methods) (Table 4) (Figure 1B, Comparison 2).
Alterations within each group could be related to the unique aspects of each condition. Changes in the control sulcus are related to general anatomy, RHI sulcal changes are related to repetitive head trauma induced damage, and CTE sulcal changes were related to p-tau deposition. Within controls, we observed 671 genes had an FDR adjusted p < 0.1. Of the FDR corrected genes, 579 were increased in the sulcus and 92 were decreased in the sulcus compared to the crest (Figure 4A). For cases in the RHI group, 1926 genes were found to have a nominal p < 0.05. However, only 1 gene, MANEA, reached FDR corrected significance. Of the nominally significant genes, 1127 were increased and 799 were decreased in the sulcus compared to the crest (Figure 4B). In subjects with CTE, 1147 genes reached FDR adjusted p < 0.1. 603 genes were elevated in the sulcus and 433 genes were decreased in the sulcus compared to the crest (Figure 4C). The top 10 increased and decreased genes by log2 fold change were annotated in each plot. A full list of sulcus vs. crest gene changes can be found in Supplementary File 2.
Figure 4. Genes that are differentially expressed in the sulcus compared to the crestThe sulcus was directly compared to the crest across (A) Control, (B) RHI, and (C) CTE and sulcal specific DE genes were calculated. Each dot represents one gene. Dots that are red represent genes that met FDR adjusted p < 0.1 significance. The top 10 positive and negatively enriched genes were annotated. GSEA analysis clarifies biological response processes that are occurring in the sulcus in each condition To further explore how the sulcal specific response contributed to the larger biological response, GSEA was performed using the genes that were significantly altered in the sulcus compared to the crest. Using FDR adjusted p < 0.1, 301 significant processes were observed in controls, 78 in RHI, and 129 in CTE. The observed processes represented biological changes specific to the sulcus compared to the crest in each relative group. We then wanted to compare enrichment of each biological process across groups and determine if there were similar or different directions of effect (Figure 5). For added clarity and biological relevance, each process was assigned to 1 of 11 greater biological categories similar to Figure 3. A heatmap was then created using processes that were significantly enriched in at least one category (Figure 5). The immune category had 51 (13.7%) processes. The majority of processes were observed to be elevated in the sulcus compared to the crest across all 3 sample groups. However, among the CTE group there was an increase in the number of processes that had a negative enrichment in the sulcus compare to the crest (Figure 5A). Next, there were 30 (8.0%) transcription/translation processes, the majority of which were positively enriched in the sulcus compared to the crest (Figure 5B). The category with the most assigned processes was the neuronal category with 99 (26.5%) processes. Almost all of the neuronal processes demonstrated a negative enrichment in the sulcus. Of the few processes that did have a positive enrichment, there was a trend towards decreasing after RHI and during CTE (Figure 4C). Next, there were 9 (2.4%) cytoskeletal processes. The ECM category had 34 (6.4%) processes. All the processes were positively enriched in control cases. However, there was a possible progressive increase in negatively enriched processes through RHI and CTE (Figure 5E). Mitochondria had 24 (6.4%) associated processes. The RHI group only had 1 negatively enriched pathway while control and CTE had a mixed split of positive and negative processes (Figure 5F). The signaling category had 56 (15.0%) processes and was evenly split between positive and negative enrichment (Figure 5G). The vascular category had 14 (8.0%) processes. The control group was entirely positively enriched in the sulcus but there was a switch to more negatively associated processes in RHI and CTE (Figure 5H). The endocytosis/exocytosis category had 13 (3.5%) processes and was primarily negatively enriched in the sulcus (Figure 5I). The protein processing category had 16 (4.3%) processes and was a mix of positive and negative processes. There appeared to be an increase in negative processes associated with CTE (Figure 5J). Finally, the 27 processes that did not fit into the previous 10 categories were placed into “other” (Figure 5K). Overall, throughout all 11 categories, there was a high agreement in the direction of effect in all cases. 73 (21.2%) processes did not have the same direction of effect.
Figure 5. Heatmap of sulcal specific GSEA biological processesUsing the genes that were significantly altered in the sulcus compared to the crest, we used GSEA to identify biological processes. Each process was then assigned to a greater overall biological category: A) Immune, B) Transcriptional/translational, C) Neuronal, D) Cytoskeletal, E) Extracellular matrix, F) Mitochondria/metabolism, G) Signaling, H) Vascular, I) Endocytosis/exocytosis, J) Protein processing, K) Other. [Please click on the figure to download a high-resolution version] Next, using the ratio that was created by directly comparing the differentially expressed genes in the sulcus to crest within each group, we compared how those ratios changed in RHI and CTE relative to control (Figure 1C, Comparison 3). This analysis provided additional information into how genes which were increased in the sulcus compared to the crest change after exposure to head trauma and p-tau pathology. 87 processes were found to be significantly altered in RHI vs Control, and 127 processes were altered in CTE vs Control. The same 11 greater biological processes were used to group GSEA processes and all processes that were significant for at least one of the two comparisons were included in a heatmap (Figure 6A-K). There were 54 immune processes altered (Figure 6A). This was the largest number of altered processes compared to the other 10 categories. The majority of immune processes were observed to be decreased compared to controls. Transcription/translation was the second largest with 46 processes being altered (Figure 6B). The neuronal category had 31 processes and the majority were observed to be increased compared to control (Figure 6C). Cytoskeletal had 16 processes and had more processes that decreased compared to increased (Figure 6D). ECM had 25 processes and all of them were decreased compared to control (Figure 6E). Mitochondria had 21 processes and the majority were increased compared to controls (Figure 6F). Signaling had 44, with the majority being decreased compared to control (Figure 6G). Vascular had 16 processes with all but one observed to decrease compared to controls (Figure 6H). Endocytosis/exocytosis had 9 processes with an even split of changes (Figure 6I). Protein processing had 22 processes with an even split of changes (Figure 6J). Finally, the 12 processes that didn’t fall in the previous 10 categories were assigned to an “other” category and were all decreased compared to control (Figure 6K). Overall, RHI and CTE had similar changes compared to controls. For RHI-control and CTE-control comparisons that did not have the same direction of effect, CTE-control comparisons always had reduced enrichment of those processes.
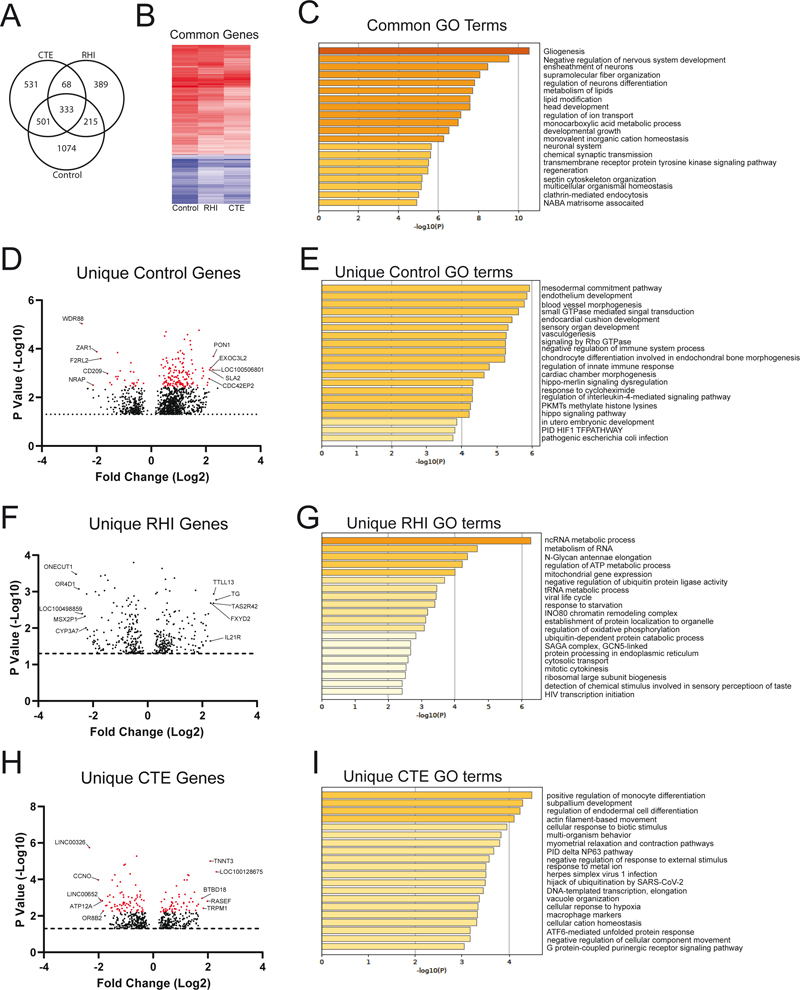
Figure 6. Heatmap of sulcal specific GSEA biological processes that change in RHI and CTE compared to controlThe RHI and CTE GSEA processes that were found to be specifically altered in the sulcus were compared to the control. Each process was again assigned to a greater overall biological category: A) Immune, B) Transcriptional/translational, C) Neuronal, D) Cytoskeletal, E) Extracellular matrix, F) Mitochondria/metabolism, G) Signaling, H) Vascular, I) Endocytosis/exocytosis, J) Protein processing, K) Other. [Please click on the figure to download a high-resolution version] Common and Unique genes found in the sulcus across disease groups Finally, using the differentially expressed gene results comparing the sulcus to the crest (Figure 1B, Comparison 2), we wanted to examine if there were any sulcal specific alterations that were unique or common across each group (Figure 7). First, we examined what sulcal specific genes were commonly altered in all three groups. 333 genes were differentially expressed in the sulcus compared with the crest among all three groups (Figure 7A). 331 of 333 genes had the same direction of effect among all three diagnostic groups (Figure 7B). Only 2 genes, KIAA2022 (up in RHI) and PLEKHH2 (down in RHI), had a conflicting direction of effect in RHI compared to control and CTE. To better understand the biological relevance of those common genes, gene ontology (GO) analysis was performed (Figure 7C). The most highly significant GO term of the common genes was “gliogenesis”, suggesting an altered glial response is an innate feature of the sulcus compared to other brain regions. When examining the rest of the top 20 significant processes, many processes were observed to encompass development, homeostasis, and organization. We then wanted to determine which differentially expressed genes in the sulcus compared with the crest were unique to each diagnostic group. A stringent criteria was used to avoid including any gene that was significant in one group but also had p values close to significant in the other two suggesting they could be common genes if power were increased. Therefore, only genes that had a nominal p < 0.05 in one group and a p > 0.25 in all others were marked to be “uniquely” altered. 1074 genes were found to be uniquely altered in the control sulcus with 806 being increased and 268 being decreased compared to the crest (Figure 7D). 168 genes had an FDR < 0.1. When observing the GO processes of the unique control genes, developmental and immune regulation processes appeared to be the most prominent (Figure 7E). 389 genes were found to be unique in the RHI group. 192 genes were increased, and 197 genes were decreased in the RHI sulcus (Figure 7F). No RHI unique gene met FDR corrected significance. Using GO analysis, it was observed that many of the processes were metabolic or transcriptionally related (Figure 6G). Finally, 531 genes were uniquely altered in the CTE sulcus. 239 genes were increased, and 292 genes were decreased in the sulcus relative to the crest (Figure 7H). 136 genes met FRD corrected significance < 0.1. GO analysis of CTE specific biological processes demonstrated immune and cell damage biological processes (Figure 7I).
Figure 7. Analysis of common and unique genes and biological process found in the sulcusUsing the genes that were found to be significantly altered in the sulcus compared to the crest, we compared which genes were common to all groups and which were unique to each condition. A stringent criteria was used to avoid including any genes that were significant in one group but also had p values close to significant others suggesting they could be common genes if power was increased. Therefore, only genes that had a p < 0.05 in one group and a p > 0.25 in all others were marked to be “uniquely” altered. A) A Venn diagram of the common and unique genes found among all three groups. B) A heatmap of the common genes showing similar directions of effect of the majority of genes. C) GO analysis of the common genes. D) A plot of the unique sulcal genes found in control cases. E) GO analysis of the unique control genes. F) A plot of the unique sulcal genes found in RHI cases. G) GO analysis of unique RHI genes. H) A plot of the unique sulcal genes found in CTE cases. I) GO analysis of unique CTE genes. In all plots, red dots denote genes that met FDR adjusted p < 0.1. GO analysis was performed using any gene that met nominal p < 0.05 significance. The top 5 positive and negative genes for each group were annotated. [Please click on the figure to download a high-resolution version] Discussion Building on previous work suggesting that altered neuroinflammation is an early event in CTE, here we have demonstrated that there is a complex inflammatory response encompassing several unique biological categories occurring specifically in the sulcus and might be directly related to CTE pathogenesis. When comparing the sulcus to the neighboring gyral crest we observed that although there was pronounced gene expression related changes present in both regions, there was a further amplified and specific sulcal alterations that might be related to head trauma and CTE pathogenesis. When comparing what genes and biological processes were altered in the sulcus compared to the crest, several higher order biological categories were observed to consistently change across control, RHI, and CTE. Immune, transcriptional/translation, ECM, and vascular categories all were positively enriched in the sulcus while neuronal and endocytosis/exocytosis processes were commonly negatively enriched in the sulcus. These findings demonstrate that even in the absence of RHI exposure or CTE pathology, the sulcus had a unique expression pattern compared to the gryal crest and might help partly explain the susceptibility to head trauma and tau deposition. When examining how the sulcal specific alterations might change in response to trauma and disease, in the RHI and CTE group there was a complex mixed increase and decrease of inflammatory processes such as astrocyte development, scavenger receptor activity, regulation of immune response, leukocyte migration, and antigen processing compared to the controls. Additionally, neuronal processes were elevated after RHI and during CTE relative to control cases, possibly suggesting injured neurons might induce compensatory mechanisms in efforts to mitigate RHI induced damage. These complex alterations could also be observed in transcriptional/translational, cytoskeletal, ECM, mitochondria/metabolism, signaling, vascular, endocytosis/exocytosis, and protein processing processes as well. Finally, we observed that each of the three groups had a set of genes and processes that were uniquely altered in the sulcus. GO analysis demonstrated that within the control group, specifically altered genes were related to development and homeostasis; within RHI, altered genes were metabolic and mitochondria related; and within CTE, altered genes were immune and inflammation related. Overall, we have built on previous work and expanded our understanding of the complex neurodegenerative response that occurs after RHI and during CTE. Since p-tau deposition in the crest is less likely in early CTE, genes that were found to be differentially expressed in both the sulcus and the crest are possibly less specific to CTE sulcal p-tau deposition and more related to a general response to head trauma. Consistent with previous reports on how repetitive head trauma affects the CNS, the most common genes that were altered in both the sulcus and crest in CTE were immune and inflammatory related (Figures 2&3). We observed that there was a significant upregulation in complement genes C4A and C4B suggesting a persistent innate immune response and activation of the complement cascade was present in the frontal cortex. Complement has a wide variety of functions, but in the context of neurodegeneration, it has been associated with increased synaptic pruning and synaptic loss [27]. It is possible the C4 activity is related to the PSD-95 synaptic protein loss that has been reported in previous CTE studies [5]. C4 has been suggested to play a role in synaptic pruning in schizophrenia [28], and C4 gene expression and protein levels have recently been linked to tau phosphatases and the development of AD suggesting a possible mechanism in the development of tau pathology [15]. Additionally, microglia related genes, such as CXCR1, were elevated in CTE, which is in agreement with previous studies examining microglia related changes in CTE [4, 7]. Astrocytic genes, GFAP and AQP1, were also found to be increased in both the sulcus and crest. It has been suggested that there is impairment of AQP4 after trauma, and it is possible that AQP1 was upregulated in compensation [31]. Interestingly, there also appeared to be a significant neutrophil response found across the sulcus and the crest. Several neutrophil related genes such as CD177, S100A8, S100A9, BPI, and LCN2, and multiple neutrophil GSEA processes were highly increased. However, it is possible that the observed neutrophil response could be related to various causes of death as opposed to contact sport exposure. Future work will be needed to closely examine if neutrophil markers could be observed using biomarkers from living individuals to better explore the possible effects of TBI and a long-term neutrophil response. Altogether, these results demonstrate that there is a prominent neuroinflammatory response found in the frontal cortex of subjects who experience repetitive head trauma. However, it is likely that changes that occur uniformly in both the sulcus and crest do not fully explain the mechanisms behind p-tau deposition and are only part of the equation of CTE mechanisms. Therefore, it was crucial that we expand our analysis to focusing on what changes could be observed uniquely in the sulcus and might directly relate to CTE pathogenesis. Interestingly, we found that multiple factors might be contributing to sulcal specific alterations. Our findings suggested that sulcal specific changes were the result of either 1) anatomy, 2) repetitive head trauma, and 3) p-tau pathology. Although the sulcus and crest are only separated by centimeters, our current result suggest that each region has a unique environment even in the absence on trauma or neurodegenerative disease. When comparing the sulcus to the crest across each group, there was high agreement in the direction of effect observed in all the processes between control, RHI, and CTE. These results provide novel information on basic neuroanatomy of how closely related tissue regions might be significantly different and result in unique neuropathologic features. To our knowledge, this is the first study to report such findings. The largest category of processes was found to be neuronal, and was downregulated in the sulcus compared to the crest. There are two possible explanations for this, either the sulcus has less neurons and supporting cells or there is a persistent neuronal impairment found in the sulcus across all groups. Regardless which is correct, it is possible that an impaired or reduced neuronal response might predispose the sulcus to elevated damage post head trauma. The next largest category of processes were immune related. Interestingly, the majority of these processes were elevated compared to the crest. This suggests that the sulcus might already be “primed” to have an exaggerated immune reaction. The primed phenotype could be consistent with having more glial present or the current glia might be predisposed to have a more severe immune reaction. Additionally, the sulcus was observed to be positively enriched in ECM and vascular processes compared to the crest, further demonstrating the sulcus is comprised of a unique environment that responds differently during pathology compared to other neighboring regions. Although it is likely the physics of head trauma induced damage and sulcal force concentration are the main driver of sulcal specific pathology [11], the findings reported here could describe additional mechanisms explaining how the sulcus is preferentially affected after repetitive head trauma [3]. Consistent with our hypothesis and previous observations, repetitive head trauma was also a significant mechanism driving sulcal specific genetic alterations. The CTE and RHI group were observed to be very similar in many genes and biological processes that were altered in the sulcus compared to controls (Figure 5). The changes that were observed to be consistent between the RHI and CTE group could be interpreted as trauma related alterations, as a common feature of both groups is they share a history of repetitive head trauma received from playing American football. It was unexpected to observe more than half of the of immune biological responses in RHI and CTE were decreased in the sulcus compared to control. Previous work had suggested that neuroinflammation was a prominent feature of head trauma and likely related to initial p-tau deposition [7]. When examining which biological processes were altered in the sulcus, it was observed that the upregulated processes were more related to foreign object recognition, antigen presentation, and lysosomal function, while the downregulated processes were related to inflammatory cytokine production and other elements related to neurodegeneration. This is consistent with our previous study demonstrating that after repetitive head trauma there was relatively unchanged numbers of total microglia, but increased levels of cells positive for CD68, a lysosomal marker that is elevated during increased phagocytosis and innate immune activation [8] Furthermore, TMEM106b genotype, which is related to lysosomal and phagocytic alterations, has been also found to relate to CTE severity [5]. Overall, the surprising downregulation of multiple immune-related processes may suggest that an impaired immune response following RHI predisposes individuals to developing initial tau pathology CTE. Future studies examining cell type specific alterations and the effects of immune-related genetic polymorphisms will be necessary to test this hypothesis. Although the CTE and RHI group did overlap for many alterations, we observed that the predominant unique RHI genes and GO processes were related to mitochondrial and metabolic activity. RHI is a group of cases that is defined by having exposure to repetitive head trauma, but no neurodegenerative pathology. Therefore, it is intriguing to observe that there was a strong metabolic deficit present even before pathology occurs. Mitochondrial activity is heavily tied to the health of the cell and any impairments could directly impact cell function. These results suggest that after a history of repetitive head trauma, mitochondrial dysfunction might be the main element driving early symptoms and pathology. Additionally, there have been several reports of mitochondria related dysfunction occurring as a consequence of head trauma [13]. It is unclear if there are a specific cell type that is most affected or what might be upstream of the mitochondrial dysfunction, but these results point towards another possible therapeutic target to treat disease before p-tau deposition even begins. Finally, we investigated the contribution of p-tau pathology to the sulcal specific genetic alterations. When directly comparing sulcal specific changes found in RHI and CTE, very few differences were observed. This was unsurprising as our CTE group was restricted to cases with low stage CTE and had minimal tau pathology. Therefore, it is likely that exposure to repetitive head trauma was a stronger driving force for sulcal specific changes than p-tau pathology in these individuals. It is also likely that many genes related to repetitive head trauma are part of the p-tau deposition mechanisms and are not necessarily unique to CTE. RHI and CTE might not be district pathologic groups, rather they exist in a spectrum. However, we were still able to identify unique genes found altered only in the CTE sulcus and therefore, likely related to p-tau deposition. The main CTE unique genes and GO processes were related to immune and cell damage further highlighting the possible connection between p-tau deposition and the immune system. It is unclear if these unique CTE genes are a consequence of p-tau deposition or act as a mechanism that, in addition to other trauma related pathways, could lead to p-tau deposition. However, these observed unique CTE biological processes are novel targets for future biomarkers studies to help identify early stage disease. There are several limitations in this study. RNA-sequencing was done on bulk tissue so no comments on the contribution of individual cells can be made at this time. Future work utilizing single cell technologies will be needed to determine the specific effects of each cell on the sulcal environment during CTE. Furthermore, it is difficult to determine the full mechanistic effect in studies using post-mortem human tissue, as we can only look at a single point in time at the end of the donor’s life. Mechanistic studies using mouse models or cell culture systems will be needed to determine if the observed genes are mechanistically related to pathologic or healthy physiologic responses. Additionally, due to the inherent variation that is present in human studies, a larger sample size is needed to identify more subtle changes. As the present study only focused on early disease, future work is also needed to compare the changes found in low stage disease to high stage CTE to determine if processes are similar. Finally, as all of the observed changes were based on mRNA, proteomic and histologic studies will be needed to verify if the results are also found at the protein level. In conclusion, these results provide clear evidence that there is a complex molecular response occurring in the sulcus compared to the gyral crest. Even without exposure to repetitive head trauma, the sulcus had a persistent immune response. In response to RHI, there was immune alteration, metabolic/mitochondria dysfunction, vascular, and ECM impairment. During CTE pathogenesis, the immune response found in low stage CTE was not observed to be tissue destruction, cell loss, overt neuroinflammation, or other neurodegenerative related responses, rather, it was more focused on antigen presentation, phagocytosis, and wound repair responses. In addition to complex immune changes, early mitochondrial, vascular, ECM, and neuronal activity are all affected during repetitive head trauma induced damage and early tau deposition, and represent possible avenues to halt CTE pathogenesis and prevent disease. These results help refine our understanding of the neuroinflammatory environment present during disease and will help guide the discovery of future therapeutic strategies and novel biomarkers. Future studies will be crucial to further dissect and study the observed biological processes provided here to better understand the full spectrum of early disease. Acknowledgements This work was supported by grant funding from: NIH (U19-AG068753, AG08122, AG054076), NIA (AG057902, AG06234, RF1AG054156, RF1AG057768), NINDS (U54NS115266, U01NS086659, and K23NS102399), National Institute of Aging Boston University AD Center (P30AG013846; supplement 0572063345-5); Department of Veterans Affairs Biorepository (BX002466) Department of Veterans Affairs Merit Award (I01-CX001038), Department of Veterans Affairs Career Development Award (BX004349), and the Nick and Lynn Buoniconti Foundation. The views, opinions, and/or findings contained in this article are those of the authors and should not be construed as an official Veterans Affairs or Department of Defense position, policy, or decision, unless so designated by other official documentation. Funders did not have a role in the design and conduct of the study; collection, management, analysis, and interpretation of the data; preparation, review, or approval of the manuscript; or decision to submit the manuscript for publication. References 1. Agus F, Crespo D, Myers RH, Labadorf A (2019) The caudate nucleus undergoes dramatic and unique transcriptional changes in human prodromal Huntington's disease brain. BMC Med Genomics 12: 137 https://doi.org/10.1186/s12920-019-0581-9 2. Alosco ML, Cherry JD, Huber BR, Tripodis Y, Baucom Z, Kowall NW et al (2020) Characterizing tau deposition in chronic traumatic encephalopathy (CTE): utility of the McKee CTE staging scheme. Acta Neuropathol 140: 495-512 https://doi.org/10.1007/s00401-020-02197-9 3. Bieniek KF, Cairns NJ, Crary JF, Dickson DW, Folkerth RD, Keene CD et al (2021) The Second NINDS/NIBIB Consensus Meeting to Define Neuropathological Criteria for the Diagnosis of Chronic Traumatic Encephalopathy. J Neuropathol Exp Neurol 80: 210-219 https://doi.org/10.1093/jnen/nlab001 4. Cherry JD, Meng G, Daley S, Xia W, Svirsky S, Alvarez VE et al (2020) CCL2 is associated with microglia and macrophage recruitment in chronic traumatic encephalopathy. J Neuroinflammation 17: 370 https://doi.org/10.1186/s12974-020-02036-4 5. Cherry JD, Mez J, Crary JF, Tripodis Y, Alvarez VE, Mahar I et al (2018) Variation in TMEM106B in chronic traumatic encephalopathy. Acta neuropathologica communications 6: 115 https://doi.org/10.1186/s40478-018-0619-9 6. Cherry JD, Olschowka JA, O'Banion MK (2014) Neuroinflammation and M2 microglia: the good, the bad, and the inflamed. J Neuroinflammation 11: 98 https://doi.org/10.1186/1742-2094-11-98 7. Cherry JD, Tripodis Y, Alvarez VE, Huber B, Kiernan PT, Daneshvar DH et al (2016) Microglial neuroinflammation contributes to tau accumulation in chronic traumatic encephalopathy. Acta Neuropathol Commun 4: 112 https://doi.org/10.1186/s40478-016-0382-8 8. Chistiakov DA, Killingsworth MC, Myasoedova VA, Orekhov AN, Bobryshev YV (2017) CD68/macrosialin: not just a histochemical marker. Lab Invest 97: 4-13 https://doi.org/10.1038/labinvest.2016.116 9. Coughlin JM, Wang Y, Minn I, Bienko N, Ambinder EB, Xu X et al (2017) Imaging of Glial Cell Activation and White Matter Integrity in Brains of Active and Recently Retired National Football League Players. JAMA Neurol 74: 67-74 https://doi.org/10.1001/jamaneurol.2016.3764 10. Coughlin JM, Wang Y, Munro CA, Ma S, Yue C, Chen S et al (2015) Neuroinflammation and brain atrophy in former NFL players: An in vivo multimodal imaging pilot study. Neurobiol Dis 74: 58-65 https://doi.org/10.1016/j.nbd.2014.10.019 11. Ghajari M, Hellyer PJ, Sharp DJ (2017) Computational modelling of traumatic brain injury predicts the location of chronic traumatic encephalopathy pathology. Brain 140: 333-343 https://doi.org/10.1093/brain/aww317 12. Hardy JA, Higgins GA (1992) Alzheimer's disease: the amyloid cascade hypothesis. Science 256: 184-185 https://doi.org/10.1126/science.1566067 13. Hiebert JB, Shen Q, Thimmesch AR, Pierce JD (2015) Traumatic brain injury and mitochondrial dysfunction. Am J Med Sci 350: 132-138 https://doi.org/10.1097/MAJ.0000000000000506 14. Huber W, Carey VJ, Gentleman R, Anders S, Carlson M, Carvalho BS et al (2015) Orchestrating high-throughput genomic analysis with Bioconductor. Nat Methods 12: 115-121 https://doi.org/10.1038/nmeth.3252 15. Jun GR, You Y, Zhu C, Meng G, Chung J, Panitch R et al (2020) Protein phosphatase 2A, complement component 4, and APOE genotype linked to Alzheimer’s disease using a systems biology approach. medRxiv: https://doi.org/10.1101/2020.11.20.20235051 16. Karran E, Mercken M, De Strooper B (2011) The amyloid cascade hypothesis for Alzheimer's disease: an appraisal for the development of therapeutics. Nat Rev Drug Discov 10: 698-712 https://doi.org/10.1038/nrd3505 17. Labadorf A, Choi SH, Myers RH (2017) Evidence for a Pan-Neurodegenerative Disease Response in Huntington's and Parkinson's Disease Expression Profiles. Front Mol Neurosci 10: 430 https://doi.org/10.3389/fnmol.2017.00430 18. Love MI, Huber W, Anders S (2014) Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15: 550 https://doi.org/10.1186/s13059-014-0550-8 19. Love S, Louis D, Ellison DW (2008) Greenfield’s Neuropathology, 2-Volume Set. CRC Press 20. Mackenzie IR, Neumann M, Bigio EH, Cairns NJ, Alafuzoff I, Kril J et al (2010) Nomenclature and nosology for neuropathologic subtypes of frontotemporal lobar degeneration: an update. Acta Neuropathol 119: 1-4 https://doi.org/10.1007/s00401-009-0612-2 21. McKee AC, Cairns NJ, Dickson DW, Folkerth RD, Dirk Keene C, Litvan I et al (2016) The first NINDS/NIBIB consensus meeting to define neuropathological criteria for the diagnosis of chronic traumatic encephalopathy. Acta Neuropathol 131: 75-86 https://doi.org/10.1007/s00401-015-1515-z 22. McKee AC, Cantu RC, Nowinski CJ, Hedley-Whyte ET, Gavett BE, Budson AE et al (2009) Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J Neuropathol Exp Neurol 68: 709-735 https://doi.org/10.1097/NEN.0b013e3181a9d503 23. McKee AC, Daneshvar DH, Alvarez VE, Stein TD (2014) The neuropathology of sport. Acta Neuropathol 127: 29-51 https://doi.org/10.1007/s00401-013-1230-6 24. McKee AC, Stern RA, Nowinski CJ, Stein TD, Alvarez VE, Daneshvar DH et al (2013) The spectrum of disease in chronic traumatic encephalopathy. Brain 136: 43-64 https://doi.org/10.1093/brain/aws307 25. McKeith IG (2006) Consensus guidelines for the clinical and pathologic diagnosis of dementia with Lewy bodies (DLB): report of the Consortium on DLB International Workshop. J Alzheimers Dis 9: 417-423. https://doi.org/10.3233/jad-2006-9s347 26. Montine TJ, Phelps CH, Beach TG, Bigio EH, Cairns NJ, Dickson DW et al (2012) National Institute on Aging-Alzheimer's Association guidelines for the neuropathologic assessment of Alzheimer's disease: a practical approach. Acta Neuropathol 123: 1-11 https://doi.org/10.1007/s00401-011-0910-3 27. Schartz ND, Tenner AJ (2020) The good, the bad, and the opportunities of the complement system in neurodegenerative disease. J Neuroinflammation 17: 354 https://doi.org/10.1186/s12974-020-02024-8 28. Sekar A, Bialas AR, de Rivera H, Davis A, Hammond TR, Kamitaki N et al (2016) Schizophrenia risk from complex variation of complement component 4. Nature 530: 177-183 https://doi.org/10.1038/nature16549 29. Sergushichev AA (2016) An algorithm for fast preranked gene set enrichment analysis using cumulative statistic calculation. BioRxiv: https://doi.org/doi.org/10.1101/060012 30. Stern RA, Daneshvar DH, Baugh CM, Seichepine DR, Montenigro PH, Riley DO et al (2013) Clinical presentation of chronic traumatic encephalopathy. Neurology 81: 1122-1129 https://doi.org/10.1212/WNL.0b013e3182a55f7f 31. Szu JI, Chaturvedi S, Patel DD, Binder DK (2020) Aquaporin-4 Dysregulation in a Controlled Cortical Impact Injury Model of Posttraumatic Epilepsy. Neuroscience 428: 140-153 https://doi.org/10.1016/j.neuroscience.2019.12.006 32. Tagge CA, Fisher AM, Minaeva OV, Gaudreau-Balderrama A, Moncaster JA, Zhang XL et al (2018) Concussion, microvascular injury, and early tauopathy in young athletes after impact head injury and an impact concussion mouse model. Brain 141: 422-458 https://doi.org/10.1093/brain/awx350 33. Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O et al (2019) Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun 10: 1523 https://doi.org/10.1038/s41467-019-09234-6
Copyright: © 2021 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |