|
|
|
Free Neuropathology 1:14 (2020) |
|
Letter |
|
Optimizing filter trap assay for the detection of aggregated alpha-synuclein in brain samples |
|
Thibauld Oullier 1*, Alice Prigent 1*, Guillaume Chapelet 1,2, Michel Neunlist 1, Franck Letournel 3, Pascal Derkinderen 1,4 |
|
1 Université de Nantes, Inserm, TENS, The Enteric Nervous System in Gut and Brain Diseases, IMAD, Nantes, France |
|
* These authors contributed equally to this work. |
|
Corresponding author: |
|
Submitted: 04 April 2020 Accepted: 26 April 2020 Copyedited by: Christian Thomas Published: 28 April 2020 |
|
Additional resources and electronic supplementary material: supplementary material |
|
Keywords: Alpha-synuclein, Parkinson’s disease, Filter trap, Dot blot, Aggregates |
|
Over the past two decades, the progress in molecular genetics and neuropathology enabled a better understanding of the pathogenesis of neurodegenerative disorders. The observation that abnormal protein accumulation is characteristic of a particular disease has led to a neuropathological classification according to the composition of the abnormal protein aggregates [1]. Several methods have been developed for the detection of these insoluble protein aggregates including, for instance, dye binding assays and immunohistochemistry with proteinase K pretreatment [1]. Filter retardation assay is a simple and rapid method which detects protein aggregates formed either in vivo or in vitro, including amyloid-beta/tau aggregates from Alzheimer’s disease (AD) [2–4] and polyglutamine expansion from Huntington’s disease [5,6]. In this assay, ‘lysis buffer’-resistant protein aggregates, but not soluble monomeric species, are retained by a cellulose acetate membrane and subsequently detected by immunoblotting. As loading control, a ‘classical’ dot blot with either polyvinylidene fluoride (PVDF) or nitrocellulose membranes is run in parallel with the same lysates. In contrast to polyglutamine expansion, tau and beta-amyloid, there are only few studies on using filter retardation assays for the detection of alpha-synuclein aggregates, which are characteristic of Parkinson’s disease (PD) and dementia with Lewy bodies (DLB) [7]. In addition, a full characterization of any filter-based assay for alpha-synuclein, including the effect of membrane composition, lysis procedure and antibodies has not yet been reported. We therefore set out the current study in which we characterize a vacuum-based 96-well format filter assay for the detection of aggregated alpha-synuclein in human brain specimens. Frozen brain samples from neuropathologically confirmed cases of DLB, progressive supranuclear palsy (PSP) and AD were obtained from the Neuropathology Department of Angers (Dr Franck Letournel). Brain samples were lysed with the “Precellys 24™” tissue homogenizer (Bertin technologies, Cat# P000669-PR240-A, Montigny-le-Bretonneux, France) followed by a brief sonication (one minute, 5 seconds on, 5 seconds off, amplitude 70%; VCX130 ultrasonic processor with a 6 mm probe, Sonics and Materials, Newton, CT, USA) either in (i) radioimmunoprecipitation assay (RIPA) buffer (Merck Millipore, Cat# 20-188, Molsheim, France) containing 2 mM orthovanadate (Sigma, Cat# S6505, Saint Quentin Fallavier, France), 1% (v/v) phosphatase inhibitor cocktail III (Sigma, Cat# P0044) and a protease inhibitors cocktail (Roche, Cat# 14424700, Meylan, France) at 4°C; (ii) ‘Sodium dodecyl sulfate (SDS) buffer’ containing 1% (w/v) SDS, 2 mM orthovanadate (Sigma, Cat# S6505) and 1% (v/v) phosphatase inhibitor cocktail III (Sigma, Cat# P0044); (iii) ‘Urea buffer’ containing 7 M urea (Sigma, Cat# T7875), 2 M thiourea (Sigma, Cat# L3771), 4% (v/v) CHAPS (Sigma, Cat# C3023), 2 mM orthovanadate (Sigma, Cat# S6505), 1% (v/v) phosphatase inhibitor cocktail III (Sigma, Cat# P0044) and a protease inhibitors cocktail (Roche, Cat# 14424700). Lysates were centrifuged at 10,000 x g for 5 minutes; supernatants were recovered and protein concentration determined using the Pierce BCA Protein Assay Kit (ThermoFisher Scientific, Cat# 23227, Saint Herblain, France) along with the Varioskan™ multimode microplate reader (ThermoFisher Scientific, Cat# VL0L00D0); samples lysed in urea buffer were diluted 5 times for compatibility with the BCA assay (https://assets.thermofisher.com/TFS-Assets/LSG/Application-Notes/TR0068-Protein-assay-compatibility.pdf). Twenty µg of these supernatants were subjected to vacuum filtration through a 96-well dot blot apparatus (Bio-Rad, Cat# 1706545, Marnes-la-Coquette, France) with 0.45 µM pore size cellulose acetate (Sterlitech, Cat# CA0453001, Kent, USA), nitrocellulose (ThermoFisher Scientific, Cat# 77010) or PVDF membranes (ThermoFisher Scientific, Cat# 88518). The resultant membranes were blocked for one hour at room temperature in Tris-buffered saline (TBS, Sigma, Cat# T5912) with 0.1% (v/v) Tween-20 (Sigma, Cat# P1379) and 5% (w/v) non-fat dry milk, then incubated overnight at 4°C with the primary antibodies listed in Table 1. Bound antibodies were detected with horseradish peroxidase-conjugated anti-rabbit (Life technologies Cat# 31460, diluted 1:5,000) or anti-mouse antibodies (Sigma, Cat# A9044, diluted 1:5,000) and visualized by enhanced chemiluminescent detection (SuperSignal™ West Pico PLUS Chemiluminescent Substrate, ThermoFisher Scientific, Cat# 34580). Oligomeric aggregates of alpha-synuclein, which were obtained by incubating monomeric recombinant alpha-synuclein (Sigma, Cat# 575001) in phosphate-buffered saline under agitation at a concentration of 10 mg/mL at 37°C for one week, served as a control [8].
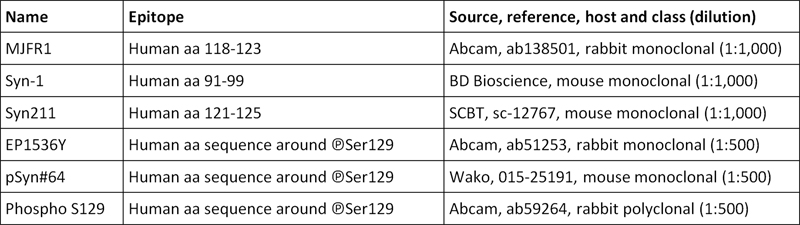
Table 1. Alpha-synuclein and phospho-alpha-synuclein antibodies used in the current study.
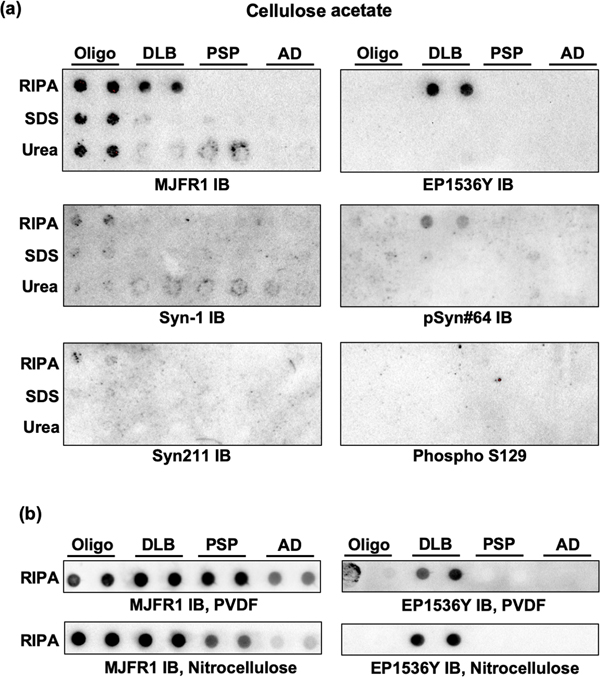
When cellulose acetate membranes were used, recombinant oligomeric alpha-synuclein and DLB lysates were barely or not detected by two of the total alpha-synuclein antibodies (Syn211 and Syn-1), regardless of the lysis procedure (Figure 1a). In contrast, MJFR1 antibody efficiently detected both oligomeric alpha-synuclein and aggregated alpha-synuclein from RIPA-lysed DLB samples (Figure 1a). No specific signal was observed with this antibody when DLB samples were lysed with SDS or urea buffers (Figure 1a). Two of the phosphospecific antibodies (EP1536Y and to a lesser extent pSyn#64) detected aggregated alpha-synuclein in RIPA-DLB lysates. No signal was observed when PSP and AD lysates were filtered and analyzed with either MJFR1 or EP1536Y antibody. These findings show that sample lysis with RIPA sample lysis together with MJFR1 and/or EP1536Y immunoblotting is the most specific and reproducible approach. We therefore used these two antibodies and RIPA-lysed samples to perform additional dot blots with PVDF or nitrocellulose membranes, that could serve as a loading control. The results show that PVDF achieved better reproducibility and specificity for the detection of total and phosphorylated alpha-synuclein than nitrocellulose membranes (Figure 1b).
Figure 1. Results from the filter trap and dot blot assays. (a) recombinant oligomeric alpha-synuclein (oligo) and samples from DLB, PSP and AD brains lysed in RIPA, SDS and Urea buffers were subjected to vacuum filtration through a 96-well dot blot apparatus with an acetate cellulose membrane; immunoblotting (IB) was then performed with 3 different antibodies to total alpha-synuclein (MJFR1, Syn-1 and Syn211) and 3 antibodies specific for the phosphorylated form of the protein (EP1536Y, pSyn#64 and Phospho S129). (b) recombinant oligomeric alpha-synuclein (Oligo) and RIPA-lysed samples from DLB, PSP and AD brains were subjected to vacuum filtration through a 96-well dot blot apparatus with a PVDF or a nitrocellulose membrane ; immunoblotting (IB) was then performed with MJFR1 or EP1536Y antibodies. Results shown are representative of 3 to 5 independent experiments.
The results presented here, although preliminary, provide the basis for reproducible and specific detection of pathological alpha-synuclein in diseased brains using a filter trap assay. We show that the selection of primary antibody and lysis buffer has important effects on the performance of the assay. As an optimized protocol, we suggest that brain homogenates should be lysed in RIPA buffer and that immunoblotting should be performed with MJFR1 for the detection of total alpha-synuclein and EP1536Y for the detection of the phosphorylated form of the protein (a step by step standard operating procedure is provided as supplementary material). Owing to the high-throughput capability of the filter trap assay, such an approach is not only potentially interesting for detecting aggregated alpha-synuclein in post mortem brain samples but also in the peripheral nervous system of living PD patients in order to develop original biomarkers of the disease [9]. References 1. Kovacs GG. Molecular Pathological Classification of Neurodegenerative Diseases: Turning towards Precision Medicine. Int J Mol Sci. 2016;17:189. 2. Bieschke J, Cohen E, Murray A, Dillin A, Kelly JW. A kinetic assessment of the C. elegans amyloid disaggregation activity enables uncoupling of disassembly and proteolysis. Protein Sci. 2009;18:2231–41. 3. Boyé-Harnasch M, Cullin C. A novel in vitro filter trap assay identifies tannic acid as an amyloid aggregation inducer for HET-s. J Biotechnol. 2006;125:222–30. 4. Chang E, Kuret J. Detection and quantification of tau aggregation using a membrane filter assay. Anal Biochem. 2008;373:330–6. 5. Novoselova TV, Margulis BA, Novoselov SS, Sapozhnikov AM, van der Spuy J, Cheetham ME, et al. Treatment with extracellular HSP70/HSC70 protein can reduce polyglutamine toxicity and aggregation. J Neurochem. 2005;94:597–606. 6. van Waarde-Verhagen MAWH, Kampinga HH. Measurement of Chaperone-Mediated Effects on Polyglutamine Protein Aggregation by the Filter Trap Assay. Methods Mol Biol. 2018;1709:59–74. 7. Recasens A, Dehay B, Bové J, Carballo-Carbajal I, Dovero S, Pérez-Villalba A, et al. Lewy body extracts from Parkinson disease brains trigger α-synuclein pathology and neurodegeneration in mice and monkeys. Ann Neurol. 2014;75:351–62. 8. Corbillé A-G, Neunlist M, Derkinderen P. Cross-linking for the analysis of α-synuclein in the enteric nervous system. J Neurochem. 2016;139:839–47. 9. Lebouvier T, Tasselli M, Paillusson S, Pouclet H, Neunlist M, Derkinderen P. Biopsable neural tissues: toward new biomarkers for Parkinson’s disease? Front Psychiatry. 2010;1:128.
Copyright: © 2020 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |