|
|
|
Free Neuropathology 1:13 (2020) |
|
Review |
|
Top ten discoveries of the year: Neurodevelopmental disorders |
|
Mara Dierssen |
|
Centre for Genomic Regulation (CRG); The Barcelona Institute of Science and Technology, and Universitat Pompeu Fabra (UPF), 08003 Barcelona, Spain |
|
Corresponding author: |
|
Submitted: 25 February 2020 Accepted: 12 April 2020 Copyedited by: Cathryn Cadwell Published: 15 April 2020 |
|
Keywords: in utero diffusion magnetic resonance imaging, in utero diffusion tensor imaging, RNA editing, Retrotransposons, Brain organoids, Single cell omics |
|
Abstract Developmental brain disorders, a highly heterogeneous group of disorders with a prevalence of around 3% of worldwide population, represent a growing medical challenge. They are characterized by impaired neurodevelopmental processes leading to deficits in cognition, social interaction, behavior and motor functioning as a result of abnormal development of brain. This can include developmental brain dysfunction, which can manifest as neuropsychiatric problems or impaired motor function, learning, language or non-verbal communication. Several of these phenotypes can often co-exist in the same patient and characterize the same disorder. Here I discuss some contributions in 2019 that are shaking our basic understanding of the pathogenesis of neurodevelopmental disorders. Recent developments in sophisticated in-utero imaging diagnostic tools have raised the possibility of imaging the fetal human brain growth, providing insights into the developing anatomy and improving diagnostics but also allowing a better understanding of antenatal pathology. On the other hand, advances in our understanding of the pathogenetic mechanisms reveal a remarkably complex molecular neuropathology involving a myriad of genetic architectures and regulatory elements that will help establish more rigorous genotype-phenotype correlations. Abbreviations 3D - three-dimensional, ADAR - adenosine deaminase acting on RNA, ASD - autism spectrum disorder, ADHD - attention deficit hyperactivity disorder, CA3 - Cornu Ammonis area 3, CEN - Central Executive Network, CNS - central nervous system, CNVs - copy number variations, DG - dentate gyrus, DLPFC - dorsolateral prefrontal cortex, DMN - Default Mode Network, DSM - The Diagnostic and Statistical Manual of Mental Disorders, DTI - diffusion tensor imaging, edQTL - editing quantitative trait loci, EEG - electroencephalography, FA - fractional anisotropy, FXS - Fragile X syndrome, GABA - γ-aminobutyric acid, GO - gene ontology, GWAS - genome-wide association study, HERV - human endogenous retroviruses, hPSCs - human pluripotent stem cells, hESCs - human embryonic stem cells, hiPSCs - human induced pluripotent stem cells, iPSCs - induced pluripotent stem cells, L1Hs - human-specific LINE-1, LINE-1 - long-interspersed nuclear element-1, MPFC - medial prefrontal cortex, MRI - magnetic resonance imaging, PGC - Psychiatric Genomics Consortium, rs-fMRI - resting-state functional MRI, scRNA-seq - single-cell RNA-sequencing, snRNA-seq - single-nucleus RNA-sequencing, TBSS - tract-based spatial statistics, TEs - transposable elements, TSA - tract-specific analysis Introduction Human brain structural and functional development occurs over a protracted period compared to many other mammals and primates (Watson et al., 2006). This coordinated development provides the architecture for the expansion of behavioral and cognitive abilities, especially rapid in the first years, but also especially vulnerable to genetic and/or environmental insults leading to developmental brain disorders. Classically those include intellectual disability, autism spectrum disorder (ASD), attention deficit hyperactivity disorder (ADHD), specific learning disorder, motor disorder, or epilepsy. However, beyond the traditional concept of neurodevelopmental disorders, studies in humans provide clear evidence that mental disorders such as schizophrenia, drug abuse, or neurodegenerative disorders such as Alzheimer’s disease also have a strong developmental component that might be identified by specific neuropathological features (Thibaut, 2018). Understanding the role that development plays in the expression of these disorders is often overlooked, but definitively needs more attention to fully understand the impact of early life events on the complex neurobiological derangement. From the clinical point of view, there has been a recent move at the diagnostic level from a categorical toward a spectrum-based view. For example, the definition of autism has been highly debated, in several revisions of the Diagnostic and Statistical Manual of Mental Disorders (DSM; DSM-III, DSM-IIIR, DSM-IV, and DSM-V) criteria (Rodgaard et al., 2019). This has led to a steady increase in the heterogeneity of some conditions which could affect the results of autism research. The recent advances in genomic medicine and the development of improved cellular models, will certainly help overcome these difficulties. On the mechanistic side, advances in directed differentiation of human induced pluripotent stem cells (hiPSCs) and other neural cell preparations (van den Ameele et al., 2014), coupled with the application of advanced histological, imaging, molecular, cellular, and genomic techniques, is providing insights into cellular and molecular processes in human neurodevelopmental disorders and also in a wide array of neurological and psychiatric disorders (Brennand et al., 2011; Lancaster et al., 2013; Mariani et al., 2015; Pasca et al., 2015). Moreover, the recent advancements in single cell functional genomic techniques have enabled comprehensive and unbiased characterization of the molecular processes in human postmortem CNS tissues and neural cell culture systems, and will allow us to explore neurodevelopmental neuropathology at a resolution that was not possible before. Even so, our understanding of the spatiotemporal landscape of the RNA species, and epigenetic features in the developing human brain, and their pathogenicity is still incomplete (Qureshi & Mehler, 2012). Finally, some findings suggest common pathophysiological mechanisms that can be considered as a continuum of developmental brain dysfunctions. These findings revealed molecular pathways that are commonly altered in different forms of developmental brain disorders, both of genetic and environmental origin and new players are starting to be recognized such as genomic regulatory elements. 1. Understanding neurodevelopmental disorders through imaging the prenatal brain One of the most crucial questions in neurodevelopmental disorders is what we can really detect from the wide range of developmental processes that take place (and can go wrong) during this critical period, and the extremely rapid pace of structural and functional brain development (Geng et al., 2017). The dynamic morphological changes the fetal brain undergoes during early development result from neurogenic events, such as neuronal proliferation, migration, axonal elongation, retraction, and myelination. In the critically sensitive mid-gestational phase of development, structural and functional assessment of the fetal brain opens a window into prenatal diagnostics and prognostics, and would help establish biomarkers for prenatal diagnoses (Batalle et al., 2017). Conventional T1 and T2 weighted sequences provide anatomic detail of the normally developing brain and can demonstrate lesions, including those associated with preterm birth, hypoxic ischemic encephalopathy, perinatal arterial stroke, infections, and congenital malformations. Specialized imaging techniques can also be used to assess cerebral vasculature (magnetic resonance angiography and venography), cerebral metabolism (magnetic resonance spectroscopy), cerebral perfusion (arterial spin labeling), and function (functional magnetic resonance imaging [MRI]) (Counsell et al., 2019). Recent advances in in utero diffusion MRI provide unique opportunities to noninvasively study the microstructure of tissue during neurodevelopment and possible mechanisms of how pathologies, maternal, or environmental factors that may interfere with brain development can be potentially detected. One important aspect is that postmortem histologic studies of early childhood cortical development are consistent with imaging studies and provide insights into the neurobiological process that underlie change observed in imaging studies (Christiaens et al., 2019). Abnormalities in processes, such as white matter and cortical connectivity in preterm babies have already been suggested to derive into late language development and impaired cognitive performance in children. in utero diffusion tensor imaging (DTI) provides new windows to monitor the emergence of the connectome. These advances have led to emerging fields of research, such as developmental pathoconnectomics (Jakab, 2019) that aim to unravel the development of disrupted brain connectivity. A wealth of quantitative tools, most of which were originally developed for the adult brain, can be applied to study the developing brain in utero and postnatally including measures of tissue microstructure obtained from diffusion MRI, morphometric studies to measure whole brain and regional tissue volumes, and automated approaches to study cortical folding. However, the differences in functional organization between the brains of infants and adults call for infant-specific functional atlases for better definition of regions of interest and interpretation of results. Thus, the creation of resources such as a DTI atlas of the fetal brain is required for reliable detection of major neuronal fiber bundle pathways and for characterization of the fetal brain reorganization in utero. Such tools are also useful for detection of normal and abnormal fetal brain development providing normative quantitative and qualitative data. The paper by Khan et al. (Khan et al., 2019) presents the first DTI atlas of the fetal brain computed from in utero diffusion-weighted images. It was built from 67 fetal DTI scans acquired from healthy fetuses each scanned at a gestational age between 21 and 39 weeks, addressing a wider gestational age range and larger number of sampled areas than prior studies. The atlas computation method represents the first comprehensive approach to compute motion-robust diffusion tensor maps from noisy in utero fetal measurements and combine individual DTI maps into a kernel-regressed template at any given gestational age which would enable statistical voxel-wise analysis, tract-based spatial statistics (TBSS), or tract-specific analysis (TSA) based on DTI data (Pecheva et al., 2017; Khan et al., 2018). The neurodevelopmental trends characterized by the atlas in the fetal brain were qualitatively and quantitatively compared with prior ex vivo and in utero studies, and with observations reported in gestational age-equivalent preterm infants. The atlas correctly detected the early presence of limbic fiber bundles followed by the appearance and maturation of projection and association fiber bundles (characterized by an age-related increase in fractal anisotropy) during late 2nd and early 3rd trimesters and the appearance and maturation of projection throughout gestation. During the 3rd trimester association fiber bundles become evident. In parallel with the appearance and maturation of fiber bundles, from 21 to 39 gestational weeks a gradual disappearance of the radial coherence of the telencephalic wall was qualitatively identified. The results presented confirm previous advanced fetal connectome imaging studies indicating increased vulnerability of the human brain during late gestation for pathologies that might lead to impaired connectome development and subsequently interfere with the development of neural substrates serving higher cognition. The atlas complements previous initiatives including recent development of detailed atlases of the fetal (Wright et al., 2015) and neonatal (Makropoulos et al., 2016) brain that allow robust automated or semi‐automated segmentation of brain regions (Makropoulos et al., 2014) and precise delineation of cortical sulcal and gyral development (Garcia et al., 2018). Together, all these tools allow characterization of the normal trajectories of fetal brain growth and creation of population centile charts (https://www.developingbrain.co.uk/fetalcentiles/ [Gousias et al., 2013]). Comparison with these typically developing growth charts therefore provides an ideal approach with which to assess, quantify, and identify deviations in regional and whole brain volumes and also in the ontogenesis, architecture, and temporal dynamics of the human brain connectome, and would lead to a more precise understanding of the etiological background of neurodevelopmental and mental disorders. These tools can be used as a reference for DTI-based studies on in utero fetal brain development, for groupwise DTI studies to investigate normal and abnormal brain development and to enable multi-modality imaging and computer-aided diagnosis from in utero DTIs. All these works certainly highlight the significant progresses we have made. However, a lack of specificity between MRI signal and neuropathological substrate is reported, as illustrated recently in one study showing that fractional anisotropy (FA) correlated with astrocyte density, a cell type typically not considered in MRI studies (Stolp et al., 2018). This indicates that the anatomical and histological significance of many “signals” in the brain remain to be investigated and further studies are needed to determine the precise cellular and molecular substrates of abnormal connectivity and gray matter microstructure observed on MRI. The new methods of three-dimensional (3D) multiscale histological imaging developed in the last years, including CLARITY (Chung & Deisseroth, 2013), MAP (Ku et al., 2016) or SHIELD (Park et al., 2018), enable rapid identification of multi-scale functional networks and interrogation of their system-wide, multifactorial interactions and thus, may help in the future for integrative and comprehensive understanding of large-scale complex biological systems. 2. Predictive value of childhood neuroimaging As discussed, in utero MRI detects fetal brain abnormalities more accurately than ultrasonography and provides additional clinical information in around half of pregnancies. However, it is less accurate when used to predict abnormal developmental outcome, although still better than ultrasonography as shown by Hart and colleagues (Hart et al., 2020). In their work published in Lancet Child and Adolescent Health, they studied the ability of in utero MRI to predict developmental outcome and whether performing postnatal neuroimaging after age 6 months changes its diagnostic accuracy. The study was performed in a cohort of children assessed with the Bayley Scales of Infant and Toddler Development, the Ages and Stages Questionnaire, or both and contrasted with ultrasonography findings. Participants' development was categorized as normal, at risk, or abnormal. However, the authors did not find statistically significant differences in infants with abnormal outcome. The authors suggest that although in utero MRI remains the optimal tool to identify fetal brain abnormalities, it is not accurate in predicting developmental outcome (Hart et al., 2020). Further work is needed to determine how the prognostic abilities of in utero MRI can be improved to identify putative infant brain markers that might be associated with neurodevelopmental disorders (Batalle et al., 2018). Instead, some works suggest that mapping the neurodevelopmental trajectories in childhood has the potential to enhance the early identification of risk. Whitfield-Gabrieli and colleagues analyzed the data from a longitudinal study of 94 children, who underwent resting-state functional MRI scans at ages 7 and 11 (Whitfield-Gabrieli et al., 2019). They explored how specific patterns of brain resting state functional connectivity change during typical development, and how these changes related to behavior. The interesting feature of this study is that children were initially recruited as typically developing, but some began to exhibit clinical symptoms over time. The aim of the study was to identify predictors of anxiety and depression behaviors in children with no familiar risk for these disorders. Specifically, the authors explored whether dysregulated top-down control mechanisms can be detected even before behavioral symptoms are evident and can predict individual children’s trajectories of attentional and internalizing problems, given that in adults with depression, anxiety, and ADHD a number of studies have shown attenuation or failure of top-down control. The strength of coupling between regions involved in top-down control and their targets can be measured with resting-state functional magnetic resonance imaging (rs-fMRI; Liu et al., 2018; Finn et al., 2015) and reliably characterizes the functional organization of the brain at a systems level (Castellanos et al., 2013). The authors focused on functional connectivity involving the Default Mode Network (DMN), a resting-state network associated with internal mentation and self-referential processing, whose key nodes include the medial prefrontal cortex (MPFC). In neurotypical adults, the DMN is negatively correlated (i.e., anticorrelated) with the Central Executive Network (CEN; Fox et al., 2005), associated with externally focused attention and goal-directed behavior, of which the dorsolateral prefrontal cortex (DLPFC) is a key node. The magnitude of the anticorrelations between the MPFC and the DLPFC is significantly correlated with superior cognitive performance such as working memory capacity (e.g., Hampson et al., 2010; Keller et al., 2015). In typically developing children, the magnitude of anticorrelations between the MPFC and DLPFC increases with age along with the improvement of top-down control mechanisms. In their study Whitfield-Gabrieli and colleagues detected individual differences in functional connectivity of the DLPFC that predicted subsequent appearance of symptoms associated with ADHD and depression. Specifically, weaker positive functional connectivity between the DLPFC and the MPFC at age 7 was associated with a decrease in ADHD symptoms by age 11, whereas weaker positive functional connectivity between the DLPFC and the subgenual anterior cingulate cortex was associated with an increase in mood-related symptoms by age 11. In fact, brain connectivity at age 7 predicted mood-related difficulties at age 11 better than baseline clinical symptoms themselves. A limitation of the work is the lack of information about which children eventually developed psychiatric disorders in this sample later on. However, these results suggest the potential utility of connectivity patterns as a biomarker of symptom trajectories. 3. The neurodevelopmental neuropathology of schizophrenia During the last year studies performed in humans have provided clear evidence that mental disorders such as schizophrenia have a strong developmental component that might be identified by specific neuropathological components. Earlier neuroimaging works reported gray matter deficits in schizophrenic patients, mostly localized in frontal and temporal lobes, which are present prior to the onset of psychosis and worsen during the first few years of illness (Glahn et al., 2008). More recently, widespread white matter decline was shown in whole-brain MRI of schizophrenia patients, identifying it as a dysconnectivity syndrome, instead of just a cortical lesion syndrome (Fornito et al., 2015). Several theories of schizophrenia suggest that structural white matter pathologies may follow developmental (McGrath et al., 2003; Murray et al., 2017), maturational (van Haren et al., 2008; French et al., 2015), and/or degenerative (Cropley et al., 2017) trajectories. Cetin-Karayumak et al. (Cetin-Karayumak et al., 2019) have addressed this question and also studied whether structural white matter pathologies vary among fiber tracts across the brain. To this aim they analyzed the largest sample of harmonized diffusion MRI data to comprehensively characterize age-related white matter neuropathology, as measured by FA. The analysis comprised diffusion scans of 600 schizophrenia patients and 492 healthy controls at different illness stages and ages (14–65 years), gathered from 13 sites. The authors cross-sectionally determined the pattern of age-related FA changes associated with schizophrenia. In whole-brain white matter, FA was up to 7% lower across the lifespan and reached peak maturation earlier in patients (27 years) than controls (33 years), and three distinct patterns of neuropathology could be identified. They detected tract-specific early developmental abnormalities in limbic fibers that do not progress over time, suggesting that limbic connections are selectively vulnerable to early developmental anomalies. Instead, long-range intra-hemispheric association tracts (including language tracts) displayed abnormal maturation with shorter maturational windows and faster declines consistent with accelerated ageing processes in schizophrenia. Finally, the authors report accelerated aging in callosal fibers that exhibited severe deficits from the outset of illness, which became more pronounced with increasing age (reaching a 10% reduction after sixth decade). This reduced anisotropy of the corpus callosum is a well-replicated diffusion imaging finding in schizophrenia, consistent across heterogeneous patient populations and maturational phases. The study makes a timely and important contribution to the field showing that white matter neuropathology in schizophrenia involves lifelong dynamic tract-specific changes. The findings support a developmental perspective, suggesting that widely distributed white matter deficits emerge early or display perturbed maturation. In addition, callosal and long-range association (but not limbic) fibers undergo accelerated aging processes. This work provides an initial benchmark for tract-specific trajectories of white matter abnormalities. 4. Disease-in-a-dish developmental neuropathology In recent years there has been a growing emphasis on developing patient-specific cellular models that can be manipulated by the experimenter to understanding the role of different factors in shaping individual brain development and functioning. Human pluripotent stem cells (hPSCs), including human embryonic stem cells (hESCs) and human induced pluripotent stem cells (hiPSCs), have been revealed as invaluable tools for modeling human disorders, especially those with complex genetic origins (Takahashi et al., 2007; Takahashi & Yamanaka, 2006). hPSCs have the potential to differentiate into any cell or tissue type. Induced pluripotent stem cells (iPSCs) reprogrammed from patient somatic cells also offer an opportunity to recapitulate disease development in relevant cell types, and they provide novel approaches for understanding disease mechanisms. Stem cells can be used to generate organoids, organ-like 3D tissue cultures containing multiple cell types that represent accessible systems for modeling organogenesis and developmental disorders (Lyon, 2019). hPSC-derived brain organoids self-assemble to form an organized architecture, composed of progenitor, neuronal and glial cell types, resembling the fetal human brain (Jo et al., 2016; Kadoshima et al., 2017; Lancaster et al., 2013). Until recently, these in vitro systems had strong limitations. First, organoids develop without the presence of normal embryonic surrounding thus lacking the developmental and patterning cues, which are essential for organ development. Additionally, most protocols depend on the ability of stem cells to self-organize into distinct brain structures which can cause inconsistency in producing the desired tissues, resulting in heterogeneity or “batch-effects” in different batches of organoids, which can vary in quality and brain regions they generate, making it difficult to identify real phenotypes. Human brain organoids are prone to high organoid-to-organoid variability (Quadrato et al., 2016; Yoon et al., 2019; Grenier et al., 2020). Furthermore, the absence of vascularization is probably responsible for the shortage of progenitor populations, making it difficult to replicate cortical plate formation. All of this has raised doubts as to whether developmental processes of the human brain can occur outside the context of embryogenesis with a degree of reproducibility that is comparable to the endogenous tissue (Jabaudon & Lancaster, 2018). The last years have been exciting for the field, because some of these limitations are starting to be overcome. Recently, Mansour et al. (Mansour et al., 2018) showed that intracerebral transplantation of brain organoids in mice results in impressive growth of blood vessels into the human tissue, with clear benefits for cell survival and maturation compared with organoids kept in vitro. Also, Real and colleagues (Real et al., 2018) have shown the potential of these methods to model human neuropathology. They transplanted human iPSC-derived cortical neurons from two persons with Down syndrome into the adult mouse cortex and observed that those consistently organized into large (up to ~100 mm3) vascularized neuron-glia territories. Down syndrome transplants showed increased synaptic stability and reduced oscillations, thus recapitulating in part the patients’ phenotypes. Finally Velasco and colleagues (Velasco et al., 2019) showed that an organoid model of the dorsal forebrain can reliably generate a rich diversity of cell types appropriate for the human cerebral cortex. Using single-cell RNA-sequencing analysis of 166,242 cells isolated from 21 individual organoids, the authors show that the organoid-to-organoid variability is comparable to that of individual endogenous brains, and 95% of the organoids derived from different stem cell lines generate a virtually indistinguishable compendium of cell types, showing consistent reproducibility in the cell types produced. The authors thus demonstrated that establishment of terminal cell identity is a highly constrained process and that reproducible developmental trajectories of cellular diversity of the CNS does not require the context of the embryo. 5. Growing networks in a dish Most of the current organoid protocols or medium formulations favor progenitor cells, thus not enabling them to mature and mimic some aspects of the human brain development such as dynamic changes in cellular populations during maturation or the formation of long-range connectivity. In a recent work, Trujillo and colleagues (Trujillo et al., 2019) developed cortical organoids that spontaneously displayed periodic and regular oscillatory network events that are dependent on glutamatergic and γ-aminobutyric acid (GABA)-ergic signaling. They could record consistent increases in electrical activity over the span of several months reflecting the formation of a spontaneous network that displayed periodic and regular oscillatory events. These nested oscillations exhibited cross-frequency coupling, subsequently transitioning to more spatiotemporally irregular patterns, resembling features observed in preterm human electroencephalography (EEG). These results suggest that the development of structured network activity in the human neocortex takes place even in the absence of external or subcortical inputs, and open opportunities for investigating and manipulating the role of network activity in the developing human cortex. Two independent groups (Cullen et al., 2019; Kirihara et al., 2019) have developed a human stem cell-derived model of cerebral tracts and have described the phenotype and connectivity of constrained 3D human axon tracts derived from brain organoids. In the work of Kirihara and colleagues they used a microfluidic device, in which two spheroids of cortical neurons derived from hiPSCs extended axons into a microchannel between the spheroids and spontaneously formed an axon fascicle, mimicking a cortico-cortical tract that connected the two spheroids reciprocally. The axon fascicle was able to communicate electrically between the spheroids with distinct response kinetics. This cerebral tract model should provide a promising platform to study the mechanisms underlying cerebral tract development and related diseases. Using their model of cerebral tracts, the authors could recapitulate the agenesis of corpus callosum (Edwards et al., 2014), by knocking down L1CAM gene (Demyanenko et al., 1999; Siegenthaler et al., 2015) in the spheroids. Axons from the L1CAM knockdown cells exhibited significantly lower ratio of axons assembled into a bundle than the control cells, suggesting that this method can be used to model developmental disease related to cerebral tracts. Microfluidic devices, involving a two-compartment system connected by narrow grooves, have also been used to reconstruct dentate gyrus (DG)–Cornu Ammonis area 3 (CA3) circuitry in which DG neurons are cultured in one compartment and CA3 neurons are cultured in the other compartment (Sarkar et al., 2018). In this system, axonal growth is allowed through the narrow grooves connecting the two compartments whereas cell migration is restricted. Rabies virus infection of the CA3 neurons permits the detection of presynaptic neurons that monosynaptically connect to the postsynaptic CA3 neurons. These engineered neural tissue connectivity models represent a first step toward potentially reconstructing brain circuits by physically replacing neuronal populations and long-range axon tracts in the brain, and might reveal how long-range connections are altered in the brains of people with neurodevelopmental disorders. 6. Genes involved in early cortical patterning are at the heart of mental comorbidity Over the past decade, genetic studies have been quite successful at identifying rare genetic variations, including inherited and de novo mutations and copy number variations (CNVs), related to specific developmental disorders. However, evidence is mounting to suggest that genetic risk variants identified among individuals with different brain disorders may converge on common genetic pathways. The remarkably complex architecture that embraces genetic mutations of distinct types (chromosomal rearrangements, copy number variants, small indels, and nucleotide substitutions) with distinct frequencies in the population (common, rare, de novo) creates difficulties in establishing rigorous genotype-phenotype correlations (Cardoso et al., 2019). Moreover, the cumulative effect of multiple common genetic variants, i.e., polygenic risk, is now being recognized as an important indicator of neurodevelopmental and psychiatric disorders (Cross-Disorder Group of the Psychiatric Genomics Consortium, 2013). Large-scale genome-wide ASD and cross-disorder association studies with enough statistical power to estimate small effects from common genetic variants are only now emerging, and they require combining data sets from multiple, large population samples. This need has been long recognized as testified by initiatives such as the Psychiatric Genomics Consortium (PGC), the largest consortium in the history of psychiatry (Sullivan et al., 2018). Now, in the largest-ever study of its kind, published in Cell, Lee et al. (Cross-Disorder Group of the Psychiatric Genomics Consortium, 2019) identified more than 100 genetic variants that affect the risk for more than one mental health condition, indicating that distinct psychiatric diseases share a common genetic structure, as shown. Using genome-wide association to analyze genetic data from 494,162 healthy controls and 232,964 individuals diagnosed with at least one of eight common psychiatric disorders, the researchers identified 109 gene variants that affect the risk for more than one psychiatric disorder. Certain disorders shared many variants, allowing the researchers to divide the conditions into three groups of genetically-related conditions: disorders characterized by compulsive behaviors (anorexia nervosa, obsessive-compulsive disorder and, to a lesser extent, Tourette syndrome); mood and psychotic disorders (bipolar disorder, major depression and schizophrenia); and early-onset neurodevelopmental disorders (ASD, ADHD and Tourette syndrome). Importantly, genes associated with multiple disorders showed increased expression beginning in the second trimester of pregnancy and play an important role in brain development. Another research team has found the first common genetic risk variants for autism robustly associated with ASD (Grove et al., 2019) in a genome-wide association meta-analysis carried out in a Danish population resource of 18,381 individuals with ASD and 27,969 controls. The meta-analysis identified five genome-wide-significant loci. Moreover, the study identified several ASD gene variants that had especially widespread influence on the risk for a number of psychiatric disorders. Concretely, the authors report seven loci shared with other traits, obtained by combining the dataset with genome-wide association study (GWAS) results from phenotypes with significantly overlapping genetic architectures (schizophrenia, major depression, and educational attainment). The researchers also uncovered, for the first time, genetic differences between the different clinical subgroups of autism. This will pinpoint genes that separate the diagnostic groups and enable more precise diagnosis, and could orient the search for behavioral, imaging and electrophysiological markers of atypical development in the infant brain. These genetic findings provide an entirely new insight into the cross-disorder biological processes, particularly relating to neuronal function and corticogenesis, and help to triangulate on families of genes active during early cortical patterning (Morgan et al., 2019; Figure 1), associated with the establishment and maintenance of neuronal connectivity (Parikshak et al., 2015) or with the foundation of inhibitory/excitatory balance in early childhood (Marin, 2012).
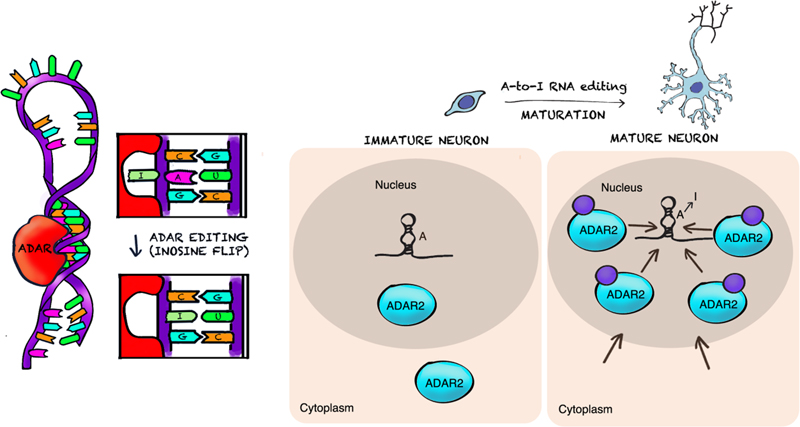
Figure 1. Left panel: Large-cohort genome-wide studies are allowing to triangulate families of genes involved in early brain maldevelopment. Middle: Early cortical patterning is at the heart of mental comorbidity. The advances in single-cell omics will help understand the pathogenesis of developmental lesions. Right panel: Critical steps of brain connectivity development have possible links to in utero MRI-detectable phenomena. For example, emergence of long-range connectivity may be detected by in-utero diffusion tensor tractography as increasing integration demonstrated using whole-brain diffusion tractography. 7. Mechanistic role of RNA editing in schizophrenia and autism neuropathology We have progressively realized in recent years that the molecular neuropathology landscape of brain disorders is much more complex than anticipated and new players, such as epigenetic regulation, alternative splicing, post-transcriptional and -translational modifications, and somatic mosaicism of DNA, not only in protein-coding sequences but also in large non-coding regions, contribute to developmental neuropathology. Transcriptomic analyses of postmortem brains have begun to elucidate some of these new mechanisms in ASD and schizophrenia. However, a crucial pathway involved in synaptic development and neuronal function, RNA editing, has not yet been studied on a genome-wide scale. RNA editing is a major RNA processing mechanism, which refers to the alteration of RNA sequences through insertion, deletion or substitution of nucleotides. It is thought to constitute one of the molecular mechanisms connecting environmental stimuli and behavioral outputs (Lapp & Hunter, 2019). Adenosine to inosine (A-to-I) editing is the most common form of RNA editing (Mallela & Nishikura, 2012; Figure 2), affecting the majority of human genes, and is highly prevalent in the brain (Liscovitch et al., 2014; Suarez et al., 2018). Editing sites in coding regions can be conserved across species and are commonly located in genes involved in neuronal function. Site-specific A-to-I RNA base conversions, carried out by adenosine deaminase acting on RNA (ADAR) enzymes, exhibit precise regional specificity in the brain and modulate complex behavior in model organisms. Examples of transcripts edited by ADAR are mRNAs encoding glutamate receptors, serotonin receptors, and potassium channels and other neuronal signaling functions in the CNS (Rosenthal & Seeburg, 2012; Meier et al., 2016; Streit & Decher, 2011). These sites are tightly and dynamically regulated throughout pre- and post-natal human cortical development (Figure 2). Most editing sites reside in non-coding regions, and particularly within sequences called transposons via RNA intermediates (retrotransposons, see below) with a possible pathophysiological role (Krestel & Meier, 2018). Aberrant RNA editing has been reported in several neurological disorders, including major depression, Alzheimer’s disease, and amyotrophic lateral sclerosis.
Figure 2. Elevated ADAR2 expression in neuronal nuclei increases A-to-I RNA editing during neuronal maturation. Left panel: ADAR enzyme acting on double-stranded RNA. ADAR RNA-binding domains act on RNA binding sites and convert adenine to inosine. Right panel: As neurons mature, A-to-I RNA editing increases gradually together with expression of importin-a4. As a result, ADAR is elevated in the nucleus leading to increased A-to-I RNA editing in mature neurons. Credits: Ilario De Toma, adapted from Behm et al., 2017 and Lorenzini et al., 2018. In ASD, recent studies in postmortem samples have revealed transcriptome dysregulation affecting neuronal and glial coding and non-coding gene expression, neuronal splicing including microexons, and microRNA targeting (Irimia et al., 2014; Salloum-Asfar et al., 2019; Gandal et al., 2018). Tran et al. (Tran et al., 2019) have now tested the largest cohort of post-mortem ASD brain samples, spanning multiple brain regions for abnormalities in RNA editing. A previous study had analyzed a few known RNA editing sites in synaptic genes and reported altered editing patterns in a small cohort of ASD cerebella (Eran et al., 2013), but it was not known whether global patterns of RNA editing contributed to the neuropathology of ASD. Now Tran and colleagues report global patterns of dysregulated RNA editing, with hypoediting across brain regions and involving many synaptic genes in ASD brains. The set of genes harboring at least one differential editing site in frontal cortex exhibited significant gene ontology (GO) enrichment for ionotropic glutamate receptor activity, glutamate gated ion channel activity, and synaptic transmission. The authors also identified a core set of down-regulated RNA editing sites, enriched in ASD susceptibility genes. Interestingly, a set of these hypoedited sites are related to Fragile X syndrome (FXS) proteins. Concretely, FMRP and FXR1P interact with ADAR and modulate A-to-I editing. The authors detected convergent dysregulated patterns of RNA editing in FXS and ASD patients, consistent with the findings that genes harboring ASD risk mutations are enriched in FMRP targets. Their findings were corroborated across multiple datasets, including dup15q cases associated with intellectual disability. Regarding schizophrenia, Breen and colleagues (Breen et al., 2019) have analyzed the global landscape and genetic regulation of RNA editing across several hundred schizophrenia and control postmortem brain samples from the dorsolateral prefrontal cortex and anterior cingulate cortex. In schizophrenia, RNA editing sites in genes encoding AMPA-type glutamate receptors and postsynaptic density proteins were less edited, whereas those encoding translation initiation machinery were more edited. These findings were cross-validated in hundreds of non-overlapping dorsolateral prefrontal cortex samples. Furthermore, ~30% of RNA editing sites associate with cis-regulatory variants (editing quantitative trait loci or edQTLs). Fine-mapping edQTLs with schizophrenia risk loci revealed co-localization of eleven edQTLs with six GWAS loci. The findings demonstrate widespread altered RNA editing in schizophrenia and its genetic regulation, and suggest a causal and mechanistic role of RNA editing in schizophrenia neuropathology. Overall, both papers provide global insights regarding RNA editing in ASD and schizophrenia pathogenesis and reveal novel mechanisms underlying these disorders. 8. Incomplete silencing of full mutation alleles in males with fragile X syndrome is associated with autistic features The paper by Tran et al. (Tran et al., 2019) convincingly demonstrated that RNA editing acts as a molecular link between FXS and ASD. FXS is caused by loss of the FMR1 product (FMRP), but also by mosaicism for active and inactive FMR1 alleles, including alleles termed premutation (55–199 CGGs). Importantly, both premutation and active full mutation (≥ 200 CGGs) alleles often express elevated levels of mRNA that are thought to be toxic. Two studies published in 2019 report that incomplete silencing of toxic full mutation RNA may be associated with autistic features in FXS males. Baker and colleagues (Baker et al., 2019) studied whether complete FMR1 mRNA silencing from full mutation alleles and/or levels of FMR1 mRNA (if present) in blood were associated with intellectual functioning and autism features in FXS. The study cohort included 98 participants (70.4% male) with FXS (full mutation-only and mosaic). FMR1 mRNA was analyzed against control FMR1 mRNA and correlated with intellectual disability and autistic features. FMR1 was completely turned off in some of the patients, while it was partially turned on in the majority of participants. Females with FXS had significantly higher levels of FMR1 mRNA that were not associated with intellectual functioning nor autistic features. In FXS males decreased levels of FMR1 mRNA were associated with decreased intellectual functioning, but not autism features. The authors show for the first time that people with a partially turned-on FMR1 had intellectual disability and more traits of ASD, whereas people with the gene completely turned off had intellectual disability and much less severe autism. The authors conclude that abnormally elevated levels of FMR1 mRNA may lead to FMR1 mRNA-related cellular “toxicity”. These findings may explain why severity of Fragile X is not the same between affected individuals. Based on this study, some months later Field et al. (Field et al., 2019) described the case of two young brothers with expanded FMR1 alleles, who were ‘high functioning’. The two brothers presented autistic features and language delay, but a higher non-verbal IQ in comparison to typical FXS. Both had low-level methylation mosaicism not detected by standard testing in blood, and their FMR1 mRNA levels were increased ~5-fold compared to typical developing controls, and significantly above the levels reported from Baker’s study described above. The authors speculate that the active unmethylated full mutation and/or premutation alleles lead to the expression of toxic expanded mRNA in some cells, in conjunction with possible reduced FMR1 mRNA and FMRP levels in other cells with FMR1 methylation. Both mechanisms may contribute to the elevated ASD symptoms. This hypothesis is also in line with the findings reported by Baker et al. showing that males who expressed full mutation FMR1 mRNA, had significantly more severe ASD symptoms compared to males who had completely silenced FMR1. 9. Transposable elements in neurodevelopmental disorders Transposable elements (TEs) constitute about half of the human genome and are becoming increasingly important to the field of neuroscience as their roles in mammalian development, immune response, and contributions to behavioral and cognitive domains continue to be uncovered (International Human Genome Sequencing Consortium, 2001; Nandi et al., 2016). Transposons are mobile DNA elements present in virtually all eukaryotes that can replicate and mobilize from one chromosomal loci to another through either a DNA or RNA intermediate (Levin & Moran, 2011). Retrotransposons are often called ‘jumping genes’, because the messenger RNA transcribed from them can undergo a process called reverse transcription to produce an identical DNA sequence that then reinserts into the genome at a different site. They parallel viruses in many ways in their structure and function, as they ensure their own survival by way of reintegration. Their propensity to self-propagate has a myriad of consequences and yet their biological significance is not well-understood. Although retrotransposons comprise about 42% of the human genome, most carry mutations that render them functionally inactive (Cordaux & Batzer, 2009). Retrotransposons are emerging as potent regulatory elements within the human genome. Transcription of those that remain functional must be prevented by protein- or RNA-based regulatory mechanisms to prevent the jumping of retrotransposons, which can cause either genetic mutations or genomic instability and might lead to cancer (Scott & Devine, 2017). Moreover, retrotransposons can be reactivated during ageing (De Cecco et al., 2013). Human Endogenous Retroviruses (HERV) and Long-Interspersed Nuclear Element-1 (LINE-1) are two main classes of retrotransposons, mobilized through a “copy and paste” mechanism. LINE-1 somatic retrotransposition has been well-demonstrated to occur in neuronal lineage, however the significance of retroelement activity to normal brain function remains uncertain. Furthermore, the contribution of these endogenous retroelements to the etiopathogenesis of neurodevelopmental disorders is a topic of recent exploration. Roughly 130 pathogenic variants caused by retrotransposon activity have been documented, but the majority of these deleterious events are isolated cases. As such, they are not part of routine clinical sequencing, and thus represent a largely unassessed category of genetic variation in many disorders. Human-specific LINE-1 (L1Hs) is the most active autonomous retrotransposon family in the human genome. Mounting evidence supports that L1Hs retrotransposition occurs postzygotically in the human brain cells, contributing to neuronal genomic diversity. In a paper in 2018 Jacob-Hirsch et al. (Jacob-Hirsch et al., 2018) already reported that the number of retrotranspositions is higher in brain tissues than that in non-brain samples and even higher in pathologic vs. normal brains. Their findings documented that L1Hs elements integrate preferentially into genes associated with neural functions and diseases. The authors propose that pre-existing retrotransposons act as “lightning rods” for novel insertions, which may safeguard from deleterious events and thus, uncontrolled retrotransposition may breach this safeguard and increase the risk of harmful mutagenesis in neurodevelopmental disorders. Zhao and colleagues (Zhao et al., 2019) profiled genome-wide L1Hs insertions among 20 postmortem tissues from Rett patients and matched controls. They identified and validated somatic L1Hs insertions in both cortical neurons and non-brain tissues, with a higher jumping activity in the brain and concluded that MeCP2 dysfunction might alter the genomic pattern of somatic L1Hs in Rett patients. Now Gardner et al. (Gardner et al., 2019) have identified retrotransposition-derived events in 9738 exome sequenced trios with developmental-affected probands. They encountered 9 de novo retrotransposons and 2 de novo gene retro-duplications. From those, 4 transposons were the likely cause of the symptoms of four patients, three of whom had not had previous diagnoses. These studies open a possibility of “diagnostic retrotransposition events” and are another step along the path to understanding the causes of developmental disorders. 10. Single-cell genomics identifies cell type–specific molecular changes in autism I have presented several efforts and advances to disentangle important elements in the molecular neuropathology of neurodevelopmental disorders. One limitation, however, is that most postmortem studies apply bulk omic approximations to a complex tissue, the brain, in which diverse human cells may contribute differently to neurodevelopmental disorders. In ASD, for example, previous bulk gene expression studies identified common genes and pathways dysregulated in the neocortex of autism patients that did not explain its clinical and genetic heterogeneity. Direct assessment of specific cell types in the brain affected by neurodevelopmental disorders has not been feasible until recent advances in techniques for isolating thousands of intact cells and efficiently sequencing each of them. Single-cell RNA-sequencing (scRNA-seq) has evolved over the past few years as a high-throughput method for transcriptome profiling of thousands of cells (Tasic, 2018) and has identified diverse cell types in many brain regions, including neocortex (Tasic et al., 2018), hypothalamus (Campbell et al., 2017), and retina (Shekhar et al., 2016). This will allow scientists to explore neurodevelopmental neuropathology at a resolution that was not possible before. An international consortium called the Human Cell Atlas (https://www.humancellatlas.org/) is an effort to identify every human cell type, where each type is located in the body, and how the cells work together to form tissues and organs. Another effort, a collaboration of 53 institutions and 60 companies across Europe, called the LifeTime Initiative (https://lifetime-fetflagship.eu/), is proposing to harness single-cell technologies to understand what happens cell by cell as tissues progress toward diseases. Transcriptomic profiling of complex tissues by scRNA-seq has unfortunately also limitations, the most important being that neurons, being complex arborizing cells, are vulnerable to mechanical dissociation and sorting. This makes scRNA-seq hard to apply to adult brain tissue to tease out individual cells from the elaborately tangled circuitry of the brain. Few studies have analyzed neurodevelopmental disorders at a single-cell resolution. A turning point came last year, when Velmeshev and colleagues (Velmeshev et al., 2019) used newer techniques to extract RNA from cell nuclei isolated from samples of brain tissue and then analyzed signature patterns of gene expression of single brain cells, including neurons and glia, from patients with autism. Single-nucleus RNA-sequencing (snRNA-seq) affords some advantages over scRNA-seq. In contrast to whole cells, nuclei are more resistant to mechanical assaults and can be isolated from frozen tissue (Krishnaswami et al., 2016), so that snRNA-seq provides less biased cellular coverage than scRNA-seq, since it suffers less cell isolation-related transcriptional artifacts (some cell types are more vulnerable to dissociation process and are thus underrepresented in the final scRNA-seq data set (Lake et al., 2016)). Velmeshev et al. applied this single-nuclei sequencing approach to snap-frozen post-mortem samples of prefrontal cortex and anterior cingulate cortex, two brain regions previously shown to be altered in patients with autism. snRNA-seq analysis of more than 100,000 cell nuclei identified a common set of changes in genes involved in synaptic communication as well as neural outgrowth and migration. Previous studies suggested convergence of ASD on specific cell types during fetal development (Parikshak et al., 2013; Willsey et al., 2013). Furthermore, in patient samples, specific sets of genes enriched in upper-layer cortico-cortical projection neurons and microglia correlated with clinical severity. They also found changes in glial cells that could impact their role in pruning and maintaining healthy neural circuits. Many of the differentially expressed genes are known to be widely expressed across the entire brain, but they appeared to be significantly altered only in projection neurons of ASD. These findings suggest that molecular changes in cell types with shared developmental lineages exhibit convergent transcriptional changes in adult ASD patients, and that the expression of synaptic and neurodevelopmental genes in layer 2/3 cortical neurons is especially affected. This implies that disturbances of gene regulatory programs during development cascade into molecular pathology in specific mature neural cell types, such as upper-layer projection neurons and microglia, which correlate with the clinical severity of ASD. One limitation of the study is the small cohort size, since the authors compared brain samples from only 15 people with autism and 16 people who died in the same age range (4-22 years) of non-neurological causes. Since almost half of the patients with autism also suffered from seizures, the researchers examined brain samples from a cohort of patients with sporadic epilepsy, to identify brain changes more likely to stem from seizures than primary ASD–associated gene expression changes. Another general limitation is that, because cells must be removed from the brain, single cell omic techniques alone do not reflect how those cells interact with their neighbors or circuit level changes. Future studies involving larger patient cohorts, including whole-exome sequencing and improved single-cell technologies, are needed to allow for precise identification of ASD-driven changes and their association with genetic variants. The authors provide an interactive web browser to interrogate their transcriptomic data: https://autism.cells.ucsc.edu. Colophon Neurodevelopmental disorders are inherently complex, involving multiple components of an intricate network. The multi-dimensional nature of the developmental disorders suggests that no unifying “cause” but instead multi-level perturbation causes complex neurodevelopmental disorders with a host of systemic comorbidities and striking heterogeneity. Many research groups are still trying to decipher the genomic complexity of disorders such as autism, and the list of autism “risk” genes grows each year. A number of new unexpected molecular players such as RNA editing and retrotransposons have been identified, and their impact on brain development is being examined. Because a deep understanding of these underlying mechanisms could prove seminal for personalized medicine, researchers have designed new cellular models with reasonable construct face and predictive validity. Of course, other factors such as the contribution of individual lifestyles, cannot be dismissed as they are fueling a large spectrum of gene-environment interactions that have a key role in the genotypes /phenotypes relationships. A more thorough understanding of the intricate processes underlying normal and abnormal human CNS development is needed to answer many fundamental questions in biology and medicine and will lead to meaningful clinical biomarkers of developmental neuropathology in humans. Funding and acknowledgements This work was supported by the Fondation Jérôme Lejeune, Paris, MINECO (SAF2016-79956-R, H2020 SC1 GO-DS21- 848077, EU (JPND HEROES (tHE cRossroad Of dEmentia Syndromes). The CRG is a Center of Excellence Severo Ochoa SEV-2016-0571. The CIBER of Rare Diseases is an initiative of the ISCIII. The laboratory of Mara Dierssen is supported by DIUE de la Generalitat de Catalunya (Grups consolidats 2017 SGR 926). We also acknowledge the support of the Spanish Ministry of Science and Innovation to the EMBL partnership, the Centro de Excelencia Severo Ochoa and the CERCA Programme/Generalitat de Catalunya. References Baker, E.K., Arpone, M., Aliaga, S.M., Bretherton, L., Kraan, C.M., Bui, M., Slater, H.R., Ling, L., Francis, D., Hunter, M.F., Elliott, J., Rogers, C., Field, M., Cohen, J., Cornish, K., Santa Maria, L., Faundes, V., Curotto, B., Morales, P., Trigo, C., Salas, I., Alliende, A.M., Amor, D.J. & Godler, D.E. (2019) Incomplete silencing of full mutation alleles in males with fragile X syndrome is associated with autistic features. Mol Autism, 10, 21. Batalle, D., Edwards, A.D. & O'Muircheartaigh, J. (2018) Annual Research Review: Not just a small adult brain: understanding later neurodevelopment through imaging the neonatal brain. J Child Psychol Psychiatry, 59, 350-371. Batalle, D., Hughes, E.J., Zhang, H., Tournier, J.D., Tusor, N., Aljabar, P., Wali, L., Alexander, D.C., Hajnal, J.V., Nosarti, C., Edwards, A.D. & Counsell, S.J. (2017) Early development of structural networks and the impact of prematurity on brain connectivity. Neuroimage, 149, 379-392. Behm, M., Wahlstedt, H., Widmark, A., Eriksson, M. & Ohman, M. (2017) Accumulation of nuclear ADAR2 regulates adenosine-to-inosine RNA editing during neuronal development. J Cell Sci, 130, 745-753. Breen, M.S., Dobbyn, A., Li, Q., Roussos, P., Hoffman, G.E., Stahl, E., Chess, A., Sklar, P., Li, J.B., Devlin, B., Buxbaum, J.D. & CommonMind, C. (2019) Global landscape and genetic regulation of RNA editing in cortical samples from individuals with schizophrenia. Nat Neurosci, 22, 1402-1412. Brennand, K.J., Simone, A., Jou, J., Gelboin-Burkhart, C., Tran, N., Sangar, S., Li, Y., Mu, Y., Chen, G., Yu, D., McCarthy, S., Sebat, J. & Gage, F.H. (2011) Modelling schizophrenia using human induced pluripotent stem cells. Nature, 473, 221-225. Campbell, J.N., Macosko, E.Z., Fenselau, H., Pers, T.H., Lyubetskaya, A., Tenen, D., Goldman, M., Verstegen, A.M., Resch, J.M., McCarroll, S.A., Rosen, E.D., Lowell, B.B. & Tsai, L.T. (2017) A molecular census of arcuate hypothalamus and median eminence cell types. Nat Neurosci, 20, 484-496. Cardoso, A.R., Lopes-Marques, M., Silva, R.M., Serrano, C., Amorim, A., Prata, M.J. & Azevedo, L. (2019) Essential genetic findings in neurodevelopmental disorders. Hum Genomics, 13, 31. Castellanos, F.X., Di Martino, A., Craddock, R.C., Mehta, A.D. & Milham, M.P. (2013) Clinical applications of the functional connectome. Neuroimage, 80, 527-540. Cetin-Karayumak, S., Di Biase, M.A., Chunga, N., Reid, B., Somes, N., Lyall, A.E., Kelly, S., Solgun, B., Pasternak, O., Vangel, M., Pearlson, G., Tamminga, C., Sweeney, J.A., Clementz, B., Schretlen, D., Viher, P.V., Stegmayer, K., Walther, S., Lee, J., Crow, T., James, A., Voineskos, A., Buchanan, R.W., Szeszko, P.R., Malhotra, A.K., Hegde, R., McCarley, R., Keshavan, M., Shenton, M., Rathi, Y. & Kubicki, M. (2019) White matter abnormalities across the lifespan of schizophrenia: a harmonized multi-site diffusion MRI study. Mol Psychiatry, 10.1038/s41380-019-0509-y, online ahead of print. Christiaens, D., Slator, P.J., Cordero-Grande, L., Price, A.N., Deprez, M., Alexander, D.C., Rutherford, M., Hajnal, J.V. & Hutter, J. (2019) In utero Diffusion MRI: Challenges, Advances, and Applications. Top Magn Reson Imaging, 28, 255-264. Chung, K. & Deisseroth, K. (2013) CLARITY for mapping the nervous system. Nat Methods, 10, 508-513. Cordaux, R. & Batzer, M.A. (2009) The impact of retrotransposons on human genome evolution. Nat Rev Genet, 10, 691-703. Counsell, S.J., Arichi, T., Arulkumaran, S. & Rutherford, M.A. (2019) Fetal and neonatal neuroimaging. Handb Clin Neurol, 162, 67-103. Cropley, V.L., Klauser, P., Lenroot, R.K., Bruggemann, J., Sundram, S., Bousman, C., Pereira, A., Di Biase, M.A., Weickert, T.W., Weickert, C.S., Pantelis, C. & Zalesky, A. (2017) Accelerated Gray and White Matter Deterioration With Age in Schizophrenia. Am J Psychiatry, 174, 286-295. Cross-Disorder Group of the Psychiatric Genomics Consortium. (2013) Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat Genet, 45, 984-994. Cross-Disorder Group of the Psychiatric Genomics Consortium. (2019) Genomic Relationships, Novel Loci, and Pleiotropic Mechanisms across Eight Psychiatric Disorders. Cell, 179, 1469-1482.e11. Cullen, D.K., Gordian-Velez, W.J., Struzyna, L.A., Jgamadze, D., Lim, J., Wofford, K.L., Browne, K.D. & Chen, H.I. (2019) Bundled Three-Dimensional Human Axon Tracts Derived from Brain Organoids. iScience, 21, 57-67. De Cecco, M., Criscione, S.W., Peterson, A.L., Neretti, N., Sedivy, J.M. & Kreiling, J.A. (2013) Transposable elements become active and mobile in the genomes of aging mammalian somatic tissues. Aging (Albany NY), 5, 867-883. Demyanenko, G.P., Tsai, A.Y. & Maness, P.F. (1999) Abnormalities in neuronal process extension, hippocampal development, and the ventricular system of L1 knockout mice. J Neurosci, 19, 4907-4920. Edwards, T.J., Sherr, E.H., Barkovich, A.J. & Richards, L.J. (2014) Clinical, genetic and imaging findings identify new causes for corpus callosum development syndromes. Brain, 137, 1579-1613. Eran, A., Li, J.B., Vatalaro, K., McCarthy, J., Rahimov, F., Collins, C., Markianos, K., Margulies, D.M., Brown, E.N., Calvo, S.E., Kohane, I.S. & Kunkel, L.M. (2013) Comparative RNA editing in autistic and neurotypical cerebella. Mol Psychiatry, 18, 1041-1048. Field, M., Dudding-Byth, T., Arpone, M., Baker, E.K., Aliaga, S.M., Rogers, C., Hickerton, C., Francis, D., Phelan, D.G., Palmer, E.E., Amor, D.J., Slater, H., Bretherton, L., Ling, L. & Godler, D.E. (2019) Significantly Elevated FMR1 mRNA and Mosaicism for Methylated Premutation and Full Mutation Alleles in Two Brothers with Autism Features Referred for Fragile X Testing. Int J Mol Sci, 20, 3907. Finn, E.S., Shen, X., Scheinost, D., Rosenberg, M.D., Huang, J., Chun, M.M., Papademetris, X. & Constable, R.T. (2015) Functional connectome fingerprinting: identifying individuals using patterns of brain connectivity. Nat Neurosci, 18, 1664-1671. Fornito, A., Zalesky, A. & Breakspear, M. (2015) The connectomics of brain disorders. Nat Rev Neurosci, 16, 159-172. Fox, M.D., Snyder, A.Z., Vincent, J.L., Corbetta, M., Van Essen, D.C. & Raichle, M.E. (2005) The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc Natl Acad Sci U S A, 102, 9673-9678. French, L., Gray, C., Leonard, G., Perron, M., Pike, G.B., Richer, L., Seguin, J.R., Veillette, S., Evans, C.J., Artiges, E., Banaschewski, T., Bokde, A.W., Bromberg, U., Bruehl, R., Buchel, C., Cattrell, A., Conrod, P.J., Flor, H., Frouin, V., Gallinat, J., Garavan, H., Gowland, P., Heinz, A., Lemaitre, H., Martinot, J.L., Nees, F., Orfanos, D.P., Pangelinan, M.M., Poustka, L., Rietschel, M., Smolka, M.N., Walter, H., Whelan, R., Timpson, N.J., Schumann, G., Smith, G.D., Pausova, Z. & Paus, T. (2015) Early Cannabis Use, Polygenic Risk Score for Schizophrenia and Brain Maturation in Adolescence. JAMA Psychiatry, 72, 1002-1011. Gandal, M.J., Zhang, P., Hadjimichael, E., Walker, R.L., Chen, C., Liu, S., Won, H., van Bakel, H., Varghese, M., Wang, Y., Shieh, A.W., Haney, J., Parhami, S., Belmont, J., Kim, M., Moran Losada, P., Khan, Z., Mleczko, J., Xia, Y., Dai, R., Wang, D., Yang, Y.T., Xu, M., Fish, K., Hof, P.R., Warrell, J., Fitzgerald, D., White, K., Jaffe, A.E., Psych, E.C., Peters, M.A., Gerstein, M., Liu, C., Iakoucheva, L.M., Pinto, D. & Geschwind, D.H. (2018) Transcriptome-wide isoform-level dysregulation in ASD, schizophrenia, and bipolar disorder. Science, 362, eaat8127. Garcia, K.E., Robinson, E.C., Alexopoulos, D., Dierker, D.L., Glasser, M.F., Coalson, T.S., Ortinau, C.M., Rueckert, D., Taber, L.A., Van Essen, D.C., Rogers, C.E., Smyser, C.D. & Bayly, P.V. (2018) Dynamic patterns of cortical expansion during folding of the preterm human brain. Proc Natl Acad Sci U S A, 115, 3156-3161. Gardner, E.J., Prigmore, E., Gallone, G., Danecek, P., Samocha, K.E., Handsaker, J., Gerety, S.S., Ironfield, H., Short, P.J., Sifrim, A., Singh, T., Chandler, K.E., Clement, E., Lachlan, K.L., Prescott, K., Rosser, E., FitzPatrick, D.R., Firth, H.V. & Hurles, M.E. (2019) Contribution of retrotransposition to developmental disorders. Nat Commun, 10, 4630. Geng, X., Li, G., Lu, Z., Gao, W., Wang, L., Shen, D., Zhu, H. & Gilmore, J.H. (2017) Structural and Maturational Covariance in Early Childhood Brain Development. Cereb Cortex, 27, 1795-1807. Glahn, D.C., Laird, A.R., Ellison-Wright, I., Thelen, S.M., Robinson, J.L., Lancaster, J.L., Bullmore, E. & Fox, P.T. (2008) Meta-analysis of gray matter anomalies in schizophrenia: application of anatomic likelihood estimation and network analysis. Biol Psychiatry, 64, 774-781. Gousias, I.S., Hammers, A., Counsell, S.J., Srinivasan, L., Rutherford, M.A., Heckemann, R.A., Hajnal, J.V., Rueckert, D. & Edwards, A.D. (2013) Magnetic resonance imaging of the newborn brain: automatic segmentation of brain images into 50 anatomical regions. PLoS One, 8, e59990. Grenier, K., Kao, J. & Diamandis, P. (2020) Three-dimensional modeling of human neurodegeneration: brain organoids coming of age. Mol Psychiatry, 25, 254-274. Grove, J., Ripke, S., Als, T.D., Mattheisen, M., Walters, R.K., Won, H., Pallesen, J., Agerbo, E., Andreassen, O.A., Anney, R., Awashti, S., Belliveau, R., Bettella, F., Buxbaum, J.D., Bybjerg-Grauholm, J., Baekvad-Hansen, M., Cerrato, F., Chambert, K., Christensen, J.H., Churchhouse, C., Dellenvall, K., Demontis, D., De Rubeis, S., Devlin, B., Djurovic, S., Dumont, A.L., Goldstein, J.I., Hansen, C.S., Hauberg, M.E., Hollegaard, M.V., Hope, S., Howrigan, D.P., Huang, H., Hultman, C.M., Klei, L., Maller, J., Martin, J., Martin, A.R., Moran, J.L., Nyegaard, M., Naerland, T., Palmer, D.S., Palotie, A., Pedersen, C.B., Pedersen, M.G., dPoterba, T., Poulsen, J.B., Pourcain, B.S., Qvist, P., Rehnstrom, K., Reichenberg, A., Reichert, J., Robinson, E.B., Roeder, K., Roussos, P., Saemundsen, E., Sandin, S., Satterstrom, F.K., Davey Smith, G., Stefansson, H., Steinberg, S., Stevens, C.R., Sullivan, P.F., Turley, P., Walters, G.B., Xu, X., Autism Spectrum Disorder Working Group of the Psychiatric Genomics Consortium, BUPGEN, Major Depressive Disorder Working Group of the Psychiatric Genomics Consortium, 23andMe Research Team, Stefansson, K., Geschwind, D.H., Nordentoft, M., Hougaard, D.M., Werge, T., Mors, O., Mortensen, P.B., Neale, B.M., Daly, M.J. & Borglum, A.D. (2019) Identification of common genetic risk variants for autism spectrum disorder. Nat Genet, 51, 431-444. Hampson, M., Driesen, N., Roth, J.K., Gore, J.C. & Constable, R.T. (2010) Functional connectivity between task-positive and task-negative brain areas and its relation to working memory performance. Magn Reson Imaging, 28, 1051-1057. Hart, A.R., Embleton, N.D., Bradburn, M., Connolly, D.J.A., Mandefield, L., Mooney, C. & Griffiths, P.D. (2020) Accuracy of in-utero MRI to detect fetal brain abnormalities and prognosticate developmental outcome: postnatal follow-up of the MERIDIAN cohort. Lancet Child Adolesc Health, 4, 131-140. International Human Genome Sequencing Consortium (2001) Initial sequencing and analysis of the human genome. Nature, 409, 860-921. Irimia, M., Weatheritt, R.J., Ellis, J.D., Parikshak, N.N., Gonatopoulos-Pournatzis, T., Babor, M., Quesnel-Vallieres, M., Tapial, J., Raj, B., O'Hanlon, D., Barrios-Rodiles, M., Sternberg, M.J., Cordes, S.P., Roth, F.P., Wrana, J.L., Geschwind, D.H. & Blencowe, B.J. (2014) A highly conserved program of neuronal microexons is misregulated in autistic brains. Cell, 159, 1511-1523. Jabaudon, D. & Lancaster, M. (2018) Exploring landscapes of brain morphogenesis with organoids. Development, 145, dev172049. Jacob-Hirsch, J., Eyal, E., Knisbacher, B.A., Roth, J., Cesarkas, K., Dor, C., Farage-Barhom, S., Kunik, V., Simon, A.J., Gal, M., Yalon, M., Moshitch-Moshkovitz, S., Tearle, R., Constantini, S., Levanon, E.Y., Amariglio, N. & Rechavi, G. (2018) Whole-genome sequencing reveals principles of brain retrotransposition in neurodevelopmental disorders. Cell Res, 28, 187-203. Jakab, A. (2019) Developmental Pathoconnectomics and Advanced Fetal MRI. Top Magn Reson Imaging, 28, 275-284. Jo, J., Xiao, Y., Sun, A.X., Cukuroglu, E., Tran, H.D., Goke, J., Tan, Z.Y., Saw, T.Y., Tan, C.P., Lokman, H., Lee, Y., Kim, D., Ko, H.S., Kim, S.O., Park, J.H., Cho, N.J., Hyde, T.M., Kleinman, J.E., Shin, J.H., Weinberger, D.R., Tan, E.K., Je, H.S. & Ng, H.H. (2016) Midbrain-like Organoids from Human Pluripotent Stem Cells Contain Functional Dopaminergic and Neuromelanin-Producing Neurons. Cell Stem Cell, 19, 248-257. Kadoshima, T., Sakaguchi, H. & Eiraku, M. (2017) Generation of Various Telencephalic Regions from Human Embryonic Stem Cells in Three-Dimensional Culture. Methods Mol Biol, 1597, 1-16. Keller, J.B., Hedden, T., Thompson, T.W., Anteraper, S.A., Gabrieli, J.D. & Whitfield-Gabrieli, S. (2015) Resting-state anticorrelations between medial and lateral prefrontal cortex: association with working memory, aging, and individual differences. Cortex, 64, 271-280. Khan, S., Hashmi, J.A., Mamashli, F., Michmizos, K., Kitzbichler, M.G., Bharadwaj, H., Bekhti, Y., Ganesan, S., Garel, K.A., Whitfield-Gabrieli, S., Gollub, R.L., Kong, J., Vaina, L.M., Rana, K.D., Stufflebeam, S.M., Hamalainen, M.S. & Kenet, T. (2018) Maturation trajectories of cortical resting-state networks depend on the mediating frequency band. Neuroimage, 174, 57-68. Khan, S., Vasung, L., Marami, B., Rollins, C.K., Afacan, O., Ortinau, C.M., Yang, E., Warfield, S.K. & Gholipour, A. (2019) Fetal brain growth portrayed by a spatiotemporal diffusion tensor MRI atlas computed from in utero images. Neuroimage, 185, 593-608. Kirihara, T., Luo, Z., Chow, S.Y.A., Misawa, R., Kawada, J., Shibata, S., Khoyratee, F., Vollette, C.A., Volz, V., Levi, T., Fujii, T. & Ikeuchi, Y. (2019) A Human Induced Pluripotent Stem Cell-Derived Tissue Model of a Cerebral Tract Connecting Two Cortical Regions. iScience, 14, 301-311. Krestel, H. & Meier, J.C. (2018) RNA Editing and Retrotransposons in Neurology. Front Mol Neurosci, 11, 163. Krishnaswami, S.R., Grindberg, R.V., Novotny, M., Venepally, P., Lacar, B., Bhutani, K., Linker, S.B., Pham, S., Erwin, J.A., Miller, J.A., Hodge, R., McCarthy, J.K., Kelder, M., McCorrison, J., Aevermann, B.D., Fuertes, F.D., Scheuermann, R.H., Lee, J., Lein, E.S., Schork, N., McConnell, M.J., Gage, F.H. & Lasken, R.S. (2016) Using single nuclei for RNA-seq to capture the transcriptome of postmortem neurons. Nat Protoc, 11, 499-524. Ku, T., Swaney, J., Park, J.Y., Albanese, A., Murray, E., Cho, J.H., Park, Y.G., Mangena, V., Chen, J. & Chung, K. (2016) Multiplexed and scalable super-resolution imaging of three-dimensional protein localization in size-adjustable tissues. Nat Biotechnol, 34, 973-981. Lake, B.B., Ai, R., Kaeser, G.E., Salathia, N.S., Yung, Y.C., Liu, R., Wildberg, A., Gao, D., Fung, H.L., Chen, S., Vijayaraghavan, R., Wong, J., Chen, A., Sheng, X., Kaper, F., Shen, R., Ronaghi, M., Fan, J.B., Wang, W., Chun, J. & Zhang, K. (2016) Neuronal subtypes and diversity revealed by single-nucleus RNA sequencing of the human brain. Science, 352, 1586-1590. Lancaster, M.A., Renner, M., Martin, C.A., Wenzel, D., Bicknell, L.S., Hurles, M.E., Homfray, T., Penninger, J.M., Jackson, A.P. & Knoblich, J.A. (2013) Cerebral organoids model human brain development and microcephaly. Nature, 501, 373-379. Lapp, H.E. & Hunter, R.G. (2019) Early life exposures, neurodevelopmental disorders, and transposable elements. Neurobiol Stress, 11, 100174. Levin, H.L. & Moran, J.V. (2011) Dynamic interactions between transposable elements and their hosts. Nat Rev Genet, 12, 615-627. Liscovitch, N., Bazak, L., Levanon, E.Y. & Chechik, G. (2014) Positive correlation between ADAR expression and its targets suggests a complex regulation mediated by RNA editing in the human brain. RNA Biol, 11, 1447-1456. Liu, J., Liao, X., Xia, M. & He, Y. (2018) Chronnectome fingerprinting: Identifying individuals and predicting higher cognitive functions using dynamic brain connectivity patterns. Hum Brain Mapp, 39, 902-915. Lorenzini, I., Moore, S. & Sattler, R. (2018) RNA Editing Deficiency in Neurodegeneration. Adv Neurobiol, 20, 63-83. Lyon, L. (2019) Building brains: using brain organoids to study neural development and disease. Brain, 142, e65. Makropoulos, A., Aljabar, P., Wright, R., Huning, B., Merchant, N., Arichi, T., Tusor, N., Hajnal, J.V., Edwards, A.D., Counsell, S.J. & Rueckert, D. (2016) Regional growth and atlasing of the developing human brain. Neuroimage, 125, 456-478. Makropoulos, A., Gousias, I.S., Ledig, C., Aljabar, P., Serag, A., Hajnal, J.V., Edwards, A.D., Counsell, S.J. & Rueckert, D. (2014) Automatic whole brain MRI segmentation of the developing neonatal brain. IEEE Trans Med Imaging, 33, 1818-1831. Mallela, A. & Nishikura, K. (2012) A-to-I editing of protein coding and noncoding RNAs. Crit Rev Biochem Mol Biol, 47, 493-501. Mansour, A.A., Goncalves, J.T., Bloyd, C.W., Li, H., Fernandes, S., Quang, D., Johnston, S., Parylak, S.L., Jin, X. & Gage, F.H. (2018) An in vivo model of functional and vascularized human brain organoids. Nat Biotechnol, 36, 432-441. Mariani, J., Coppola, G., Zhang, P., Abyzov, A., Provini, L., Tomasini, L., Amenduni, M., Szekely, A., Palejev, D., Wilson, M., Gerstein, M., Grigorenko, E.L., Chawarska, K., Pelphrey, K.A., Howe, J.R. & Vaccarino, F.M. (2015) FOXG1-Dependent Dysregulation of GABA/Glutamate Neuron Differentiation in Autism Spectrum Disorders. Cell, 162, 375-390. Marin, O. (2012) Interneuron dysfunction in psychiatric disorders. Nat Rev Neurosci, 13, 107-120. McGrath, J.J., Feron, F.P., Burne, T.H., Mackay-Sim, A. & Eyles, D.W. (2003) The neurodevelopmental hypothesis of schizophrenia: a review of recent developments. Ann Med, 35, 86-93. Meier, J.C., Kankowski, S., Krestel, H. & Hetsch, F. (2016) RNA Editing-Systemic Relevance and Clue to Disease Mechanisms? Front Mol Neurosci, 9, 124. Morgan, S.E., Seidlitz, J., Whitaker, K.J., Romero-Garcia, R., Clifton, N.E., Scarpazza, C., van Amelsvoort, T., Marcelis, M., van Os, J., Donohoe, G., Mothersill, D., Corvin, A., Pocklington, A., Raznahan, A., McGuire, P., Vertes, P.E. & Bullmore, E.T. (2019) Cortical patterning of abnormal morphometric similarity in psychosis is associated with brain expression of schizophrenia-related genes. Proc Natl Acad Sci U S A, 116, 9604-9609. Murray, R.M., Bhavsar, V., Tripoli, G. & Howes, O. (2017) 30 Years on: How the Neurodevelopmental Hypothesis of Schizophrenia Morphed Into the Developmental Risk Factor Model of Psychosis. Schizophr Bull, 43, 1190-1196. Nandi, S., Chandramohan, D., Fioriti, L., Melnick, A.M., Hebert, J.M., Mason, C.E., Rajasethupathy, P. & Kandel, E.R. (2016) Roles for small noncoding RNAs in silencing of retrotransposons in the mammalian brain. Proc Natl Acad Sci U S A, 113, 12697-12702. Parikshak, N.N., Gandal, M.J. & Geschwind, D.H. (2015) Systems biology and gene networks in neurodevelopmental and neurodegenerative disorders. Nat Rev Genet, 16, 441-458. Parikshak, N.N., Luo, R., Zhang, A., Won, H., Lowe, J.K., Chandran, V., Horvath, S. & Geschwind, D.H. (2013) Integrative functional genomic analyses implicate specific molecular pathways and circuits in autism. Cell, 155, 1008-1021. Park, Y.G., Sohn, C.H., Chen, R., McCue, M., Yun, D.H., Drummond, G.T., Ku, T., Evans, N.B., Oak, H.C., Trieu, W., Choi, H., Jin, X., Lilascharoen, V., Wang, J., Truttmann, M.C., Qi, H.W., Ploegh, H.L., Golub, T.R., Chen, S.C., Frosch, M.P., Kulik, H.J., Lim, B.K. & Chung, K. (2018) Protection of tissue physicochemical properties using polyfunctional crosslinkers. Nat Biotechnol, 10.1038/nbt.4281. Pasca, A.M., Sloan, S.A., Clarke, L.E., Tian, Y., Makinson, C.D., Huber, N., Kim, C.H., Park, J.Y., O'Rourke, N.A., Nguyen, K.D., Smith, S.J., Huguenard, J.R., Geschwind, D.H., Barres, B.A. & Pasca, S.P. (2015) Functional cortical neurons and astrocytes from human pluripotent stem cells in 3D culture. Nat Methods, 12, 671-678. Pecheva, D., Yushkevich, P., Batalle, D., Hughes, E., Aljabar, P., Wurie, J., Hajnal, J.V., Edwards, A.D., Alexander, D.C., Counsell, S.J. & Zhang, H. (2017) A tract-specific approach to assessing white matter in preterm infants. Neuroimage, 157, 675-694. Quadrato, G., Brown, J. & Arlotta, P. (2016) The promises and challenges of human brain organoids as models of neuropsychiatric disease. Nat Med, 22, 1220-1228. Qureshi, I.A. & Mehler, M.F. (2012) Emerging roles of non-coding RNAs in brain evolution, development, plasticity and disease. Nat Rev Neurosci, 13, 528-541. Real, R., Peter, M., Trabalza, A., Khan, S., Smith, M.A., Dopp, J., Barnes, S.J., Momoh, A., Strano, A., Volpi, E., Knott, G., Livesey, F.J. & De Paola, V. (2018) In vivo modeling of human neuron dynamics and Down syndrome. Science, 362, eaau1810. Rodgaard, E.M., Jensen, K., Vergnes, J.N., Soulieres, I. & Mottron, L. (2019) Temporal Changes in Effect Sizes of Studies Comparing Individuals With and Without Autism: A Meta-analysis. JAMA Psychiatry, 76, 1124-1132. Rosenthal, J.J. & Seeburg, P.H. (2012) A-to-I RNA editing: effects on proteins key to neural excitability. Neuron, 74, 432-439. Salloum-Asfar, S., Satheesh, N.J. & Abdulla, S.A. (2019) Circulating miRNAs, Small but Promising Biomarkers for Autism Spectrum Disorder. Front Mol Neurosci, 12, 253. Sarkar, A., Mei, A., Paquola, A.C.M., Stern, S., Bardy, C., Klug, J.R., Kim, S., Neshat, N., Kim, H.J., Ku, M., Shokhirev, M.N., Adamowicz, D.H., Marchetto, M.C., Jappelli, R., Erwin, J.A., Padmanabhan, K., Shtrahman, M., Jin, X. & Gage, F.H. (2018) Efficient Generation of CA3 Neurons from Human Pluripotent Stem Cells Enables Modeling of Hippocampal Connectivity In Vitro. Cell Stem Cell, 22, 684-697.e9. Scott, E.C. & Devine, S.E. (2017) The Role of Somatic L1 Retrotransposition in Human Cancers. Viruses, 9, 131. Shekhar, K., Lapan, S.W., Whitney, I.E., Tran, N.M., Macosko, E.Z., Kowalczyk, M., Adiconis, X., Levin, J.Z., Nemesh, J., Goldman, M., McCarroll, S.A., Cepko, C.L., Regev, A. & Sanes, J.R. (2016) Comprehensive Classification of Retinal Bipolar Neurons by Single-Cell Transcriptomics. Cell, 166, 1308-1323.e30. Siegenthaler, D., Enneking, E.M., Moreno, E. & Pielage, J. (2015) L1CAM/Neuroglian controls the axon-axon interactions establishing layered and lobular mushroom body architecture. J Cell Biol, 208, 1003-1018. Stolp, H.B., Ball, G., So, P.W., Tournier, J.D., Jones, M., Thornton, C. & Edwards, A.D. (2018) Voxel-wise comparisons of cellular microstructure and diffusion-MRI in mouse hippocampus using 3D Bridging of Optically-clear histology with Neuroimaging Data (3D-BOND). Sci Rep, 8, 4011. Streit, A.K. & Decher, N. (2011) A-to-I RNA editing modulates the pharmacology of neuronal ion channels and receptors. Biochemistry (Mosc), 76, 890-899. Suarez, N.A., Macia, A. & Muotri, A.R. (2018) LINE-1 retrotransposons in healthy and diseased human brain. Dev Neurobiol, 78, 434-455. Sullivan, P.F., Agrawal, A., Bulik, C.M., Andreassen, O.A., Borglum, A.D., Breen, G., Cichon, S., Edenberg, H.J., Faraone, S.V., Gelernter, J., Mathews, C.A., Nievergelt, C.M., Smoller, J.W., O'Donovan, M.C. & Psychiatric Genomics, C. (2018) Psychiatric Genomics: An Update and an Agenda. Am J Psychiatry, 175, 15-27. Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka, T., Tomoda, K. & Yamanaka, S. (2007) Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell, 131, 861-872. Takahashi, K. & Yamanaka, S. (2006) Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell, 126, 663-676. Tasic, B. (2018) Single cell transcriptomics in neuroscience: cell classification and beyond. Curr Opin Neurobiol, 50, 242-249. Tasic, B., Yao, Z., Graybuck, L.T., Smith, K.A., Nguyen, T.N., Bertagnolli, D., Goldy, J., Garren, E., Economo, M.N., Viswanathan, S., Penn, O., Bakken, T., Menon, V., Miller, J., Fong, O., Hirokawa, K.E., Lathia, K., Rimorin, C., Tieu, M., Larsen, R., Casper, T., Barkan, E., Kroll, M., Parry, S., Shapovalova, N.V., Hirschstein, D., Pendergraft, J., Sullivan, H.A., Kim, T.K., Szafer, A., Dee, N., Groblewski, P., Wickersham, I., Cetin, A., Harris, J.A., Levi, B.P., Sunkin, S.M., Madisen, L., Daigle, T.L., Looger, L., Bernard, A., Phillips, J., Lein, E., Hawrylycz, M., Svoboda, K., Jones, A.R., Koch, C. & Zeng, H. (2018) Shared and distinct transcriptomic cell types across neocortical areas. Nature, 563, 72-78. Thibaut, F. (2018) Psychiatric disorders: neurodevelopmental disorders, neurodegenerative disorders, or both? Dialogues Clin Neurosci, 20, 251-252. Tran, S.S., Jun, H.I., Bahn, J.H., Azghadi, A., Ramaswami, G., Van Nostrand, E.L., Nguyen, T.B., Hsiao, Y.E., Lee, C., Pratt, G.A., Martinez-Cerdeno, V., Hagerman, R.J., Yeo, G.W., Geschwind, D.H. & Xiao, X. (2019) Widespread RNA editing dysregulation in brains from autistic individuals. Nat Neurosci, 22, 25-36. Trujillo, C.A., Gao, R., Negraes, P.D., Gu, J., Buchanan, J., Preissl, S., Wang, A., Wu, W., Haddad, G.G., Chaim, I.A., Domissy, A., Vandenberghe, M., Devor, A., Yeo, G.W., Voytek, B. & Muotri, A.R. (2019) Complex Oscillatory Waves Emerging from Cortical Organoids Model Early Human Brain Network Development. Cell Stem Cell, 25, 558-569.e7. van den Ameele, J., Tiberi, L., Vanderhaeghen, P. & Espuny-Camacho, I. (2014) Thinking out of the dish: what to learn about cortical development using pluripotent stem cells. Trends Neurosci, 37, 334-342. van Haren, N.E., Hulshoff Pol, H.E., Schnack, H.G., Cahn, W., Brans, R., Carati, I., Rais, M. & Kahn, R.S. (2008) Progressive brain volume loss in schizophrenia over the course of the illness: evidence of maturational abnormalities in early adulthood. Biol Psychiatry, 63, 106-113. Velasco, S., Kedaigle, A.J., Simmons, S.K., Nash, A., Rocha, M., Quadrato, G., Paulsen, B., Nguyen, L., Adiconis, X., Regev, A., Levin, J.Z. & Arlotta, P. (2019) Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature, 570, 523-527. Velmeshev, D., Schirmer, L., Jung, D., Haeussler, M., Perez, Y., Mayer, S., Bhaduri, A., Goyal, N., Rowitch, D.H. & Kriegstein, A.R. (2019) Single-cell genomics identifies cell type-specific molecular changes in autism. Science, 364, 685-689. Watson, R.E., Desesso, J.M., Hurtt, M.E. & Cappon, G.D. (2006) Postnatal growth and morphological development of the brain: a species comparison. Birth Defects Res B Dev Reprod Toxicol, 77, 471-484. Whitfield-Gabrieli, S., Wendelken, C., Nieto-Castanon, A., Bailey, S.K., Anteraper, S.A., Lee, Y.J., Chai, X.Q., Hirshfeld-Becker, D.R., Biederman, J., Cutting, L.E. & Bunge, S.A. (2019) Association of Intrinsic Brain Architecture With Changes in Attentional and Mood Symptoms During Development. JAMA Psychiatry, 77, 378-386. Willsey, A.J., Sanders, S.J., Li, M., Dong, S., Tebbenkamp, A.T., Muhle, R.A., Reilly, S.K., Lin, L., Fertuzinhos, S., Miller, J.A., Murtha, M.T., Bichsel, C., Niu, W., Cotney, J., Ercan-Sencicek, A.G., Gockley, J., Gupta, A.R., Han, W., He, X., Hoffman, E.J., Klei, L., Lei, J., Liu, W., Liu, L., Lu, C., Xu, X., Zhu, Y., Mane, S.M., Lein, E.S., Wei, L., Noonan, J.P., Roeder, K., Devlin, B., Sestan, N. & State, M.W. (2013) Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell, 155, 997-1007. Wright, R., Makropoulos, A., Kyriakopoulou, V., Patkee, P.A., Koch, L.M., Rutherford, M.A., Hajnal, J.V., Rueckert, D. & Aljabar, P. (2015) Construction of a fetal spatio-temporal cortical surface atlas from in utero MRI: Application of spectral surface matching. Neuroimage, 120, 467-480. Yoon, S.J., Elahi, L.S., Pasca, A.M., Marton, R.M., Gordon, A., Revah, O., Miura, Y., Walczak, E.M., Holdgate, G.M., Fan, H.C., Huguenard, J.R., Geschwind, D.H. & Pasca, S.P. (2019) Reliability of human cortical organoid generation. Nat Methods, 16, 75-78. Zhao, B., Wu, Q., Ye, A.Y., Guo, J., Zheng, X., Yang, X., Yan, L., Liu, Q.R., Hyde, T.M., Wei, L. & Huang, A.Y. (2019) Somatic LINE-1 retrotransposition in cortical neurons and non-brain tissues of Rett patients and healthy individuals. PLoS Genet, 15, e1008043.
Copyright: © 2020 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |